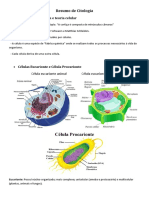
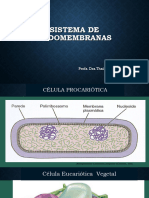
Você também pode gostar
- Sintese e Secreção de MacromoléculasDocumento7 páginasSintese e Secreção de MacromoléculasPablo MonticeliAinda não há avaliações
- As Organelas Na Síntese e Degradação Das MoléculasDocumento8 páginasAs Organelas Na Síntese e Degradação Das MoléculasSamuel LemosAinda não há avaliações
- HISTOLOGIADocumento18 páginasHISTOLOGIARebecca SicariAinda não há avaliações
- Anotações BioquímicaDocumento10 páginasAnotações BioquímicaAnderson Souza de AssisAinda não há avaliações
- Resumo: EndomembranasDocumento5 páginasResumo: EndomembranasBeatriz SantosAinda não há avaliações
- Ciclo Da Ureia1Documento6 páginasCiclo Da Ureia1HENRIQUEAinda não há avaliações
- Sistema de EndomembranasDocumento37 páginasSistema de EndomembranasRose Meire Costa BrancalhãoAinda não há avaliações
- Aula 051599763710Documento11 páginasAula 051599763710SGI Gestão IntegradoAinda não há avaliações
- BiocelDocumento10 páginasBiocelPatriciaAinda não há avaliações
- Aula 5 - BiocelDocumento13 páginasAula 5 - Biocelliz de andradeAinda não há avaliações
- O transporte entre citosol e núcleoDocumento12 páginasO transporte entre citosol e núcleoMatheus Correa0% (1)
- Resumo da estrutura e função da célulaDocumento13 páginasResumo da estrutura e função da célulaLuciana Taliano50% (2)
- Sistema de endomembranas: componentes y funcionesDocumento9 páginasSistema de endomembranas: componentes y funcionesEdgardo Andrés Bilbao MuñozAinda não há avaliações
- Organelas e transporte de proteínasDocumento14 páginasOrganelas e transporte de proteínasRapha BuenoAinda não há avaliações
- RER, Golgi, Lisossomas, Mitocôndrias e CloroplastosDocumento4 páginasRER, Golgi, Lisossomas, Mitocôndrias e CloroplastosTiago AzevedoAinda não há avaliações
- REL e RERDocumento12 páginasREL e RERAmanda FariaAinda não há avaliações
- Apostila - MitocondriasDocumento3 páginasApostila - Mitocondriastamarauescbiomed931Ainda não há avaliações
- Processos celulares e biomoléculasDocumento16 páginasProcessos celulares e biomoléculasjoanaAinda não há avaliações
- Cópia de 05 Síntese e Secreção - NewDocumento72 páginasCópia de 05 Síntese e Secreção - NewEdson AguiarAinda não há avaliações
- Ribossomos RER REL tradução proteínasDocumento8 páginasRibossomos RER REL tradução proteínasAndre Luiz do NascimentoAinda não há avaliações
- Aula 3 - RE e GolgiDocumento42 páginasAula 3 - RE e GolgiCamila HerculanoAinda não há avaliações
- Bases Biológicas Do Comportamento Aula 1Documento27 páginasBases Biológicas Do Comportamento Aula 1Tharick Matheus - Administrativo DFAinda não há avaliações
- Resumo Biologia Celular P2Documento17 páginasResumo Biologia Celular P2Ana Beatriz Rodrigues DestefanoAinda não há avaliações
- Retculo Endoplasmtico e GolgiDocumento24 páginasRetculo Endoplasmtico e GolgiCamila MendesAinda não há avaliações
- Sist - ENDOMEMBRANA 2023 - CBiomedDocumento46 páginasSist - ENDOMEMBRANA 2023 - CBiomedavenidadepirocaAinda não há avaliações
- A Síntese Proteica No Retículo Endoplasmático RugosoDocumento1 páginaA Síntese Proteica No Retículo Endoplasmático RugosoHeloise PereiraAinda não há avaliações
- Atmr 09 BiologiaDocumento13 páginasAtmr 09 Biologiapd536121Ainda não há avaliações
- Síntese proteica em 3 etapasDocumento11 páginasSíntese proteica em 3 etapasJoão SilvaAinda não há avaliações
- Biologia 11 AnoDocumento24 páginasBiologia 11 AnoIsabel Da SilvaAinda não há avaliações
- Transporte de Proteínas para Organelas por Sinais de DistribuiçãoDocumento5 páginasTransporte de Proteínas para Organelas por Sinais de DistribuiçãoMacklecio OliveiraAinda não há avaliações
- O núcleo celular: estrutura, função e componentesDocumento4 páginasO núcleo celular: estrutura, função e componentesEduardo GonçalvesAinda não há avaliações
- ENDOMEMBRANASDocumento10 páginasENDOMEMBRANASYuri AlmeidaAinda não há avaliações
- Ficha 8-BIOQDocumento8 páginasFicha 8-BIOQcassimo abilio santosAinda não há avaliações
- Células: estrutura, função e processosDocumento8 páginasCélulas: estrutura, função e processosDenise CorreiaAinda não há avaliações
- Via Secretória e OrganelosDocumento12 páginasVia Secretória e Organelosjoaopinheiro88Ainda não há avaliações
- Organização e estrutura celularDocumento43 páginasOrganização e estrutura celularMárcio Muniz100% (1)
- Cuestionario Biología - Microestructuras y procesos celularesDocumento8 páginasCuestionario Biología - Microestructuras y procesos celularesFabiana CavalcantiAinda não há avaliações
- Morfologia e Funções das OrganelasDocumento4 páginasMorfologia e Funções das OrganelasAna Carolina FernandesAinda não há avaliações
- Resumo 10-Nucleo-2009Documento8 páginasResumo 10-Nucleo-2009Isabela LiaAinda não há avaliações
- Sistema de EndomembranasDocumento26 páginasSistema de Endomembranascarmita geronimo garciaAinda não há avaliações
- Organelos e transporte de proteínas nas células eucarióticasDocumento31 páginasOrganelos e transporte de proteínas nas células eucarióticasCarolina MoreiraAinda não há avaliações
- ProteinasDocumento11 páginasProteinasLuis Mateus Camelo França MartinsAinda não há avaliações
- Roteiro P3 - Semestre 2021Documento23 páginasRoteiro P3 - Semestre 2021Ediane RubasAinda não há avaliações
- 1_4931614829476577583Documento9 páginas1_4931614829476577583Saix AbelAinda não há avaliações
- BG 2Documento5 páginasBG 2Ana RodriguesAinda não há avaliações
- Complexo de Golgi e LisossomosDocumento9 páginasComplexo de Golgi e LisossomosJúlia MottaAinda não há avaliações
- Problema 1Documento13 páginasProblema 1Moises Popp de OliveiraAinda não há avaliações
- Estudo Dirigido - P1 - Neurobiologia MolceularDocumento3 páginasEstudo Dirigido - P1 - Neurobiologia Molceulargabi.amota3Ainda não há avaliações
- Aula 14-15-16 - Sistemas de Endomembranas e Transporte CelularDocumento65 páginasAula 14-15-16 - Sistemas de Endomembranas e Transporte CelularAlejandro Morales GomezAinda não há avaliações
- Complexo de Golgi e LisossomosDocumento105 páginasComplexo de Golgi e LisossomosBruna SerraAinda não há avaliações
- Roteiro SinteseDocumento4 páginasRoteiro SintesePablo MonticeliAinda não há avaliações
- Ácidos Nucleicos, Síntese Proteica e Ciclo Celular - Biologia 11Documento8 páginasÁcidos Nucleicos, Síntese Proteica e Ciclo Celular - Biologia 11Sofia Vasconcelos Freire e CoutinhoAinda não há avaliações
- p2 Funbio 1Documento6 páginasp2 Funbio 1A MeloAinda não há avaliações
- Características compartilhadas por célulasDocumento53 páginasCaracterísticas compartilhadas por célulasMario OliveiraAinda não há avaliações
- Bioquímica I: Síntese de Proteínas, Estrutura e PurificaçãoDocumento3 páginasBioquímica I: Síntese de Proteínas, Estrutura e PurificaçãoAna FerreiraAinda não há avaliações
- Aula 4 - Endocitose e Digestao CelularDocumento46 páginasAula 4 - Endocitose e Digestao CelularCamila HerculanoAinda não há avaliações
- Endossomos MayraaDocumento29 páginasEndossomos MayraaRicardo JuperAinda não há avaliações
- Noções básicas para utilização de calculadora científicaDocumento16 páginasNoções básicas para utilização de calculadora científicaGaius Cassius LonginusAinda não há avaliações
- Cetamina em PsiquiatriaDocumento20 páginasCetamina em PsiquiatriaMilton Mendonça Junior100% (1)
- Autorização e Conferencia e Aprovação de Projetos QD 18 LT 33 CapriDocumento5 páginasAutorização e Conferencia e Aprovação de Projetos QD 18 LT 33 Caprifrancielly.agfAinda não há avaliações
- Validação da Actigrafia no Estudo do SonoDocumento54 páginasValidação da Actigrafia no Estudo do SonoAlexPsrAinda não há avaliações
- PNL em VendasDocumento4 páginasPNL em VendasRodney Formaggini100% (2)
- Mapa Astral Digitalizado - Walter SallesDocumento1 páginaMapa Astral Digitalizado - Walter SalleswaltersallesAinda não há avaliações
- Mapa Curtir A Natureza GUARAREMADocumento2 páginasMapa Curtir A Natureza GUARAREMAJuan Carlos JcAinda não há avaliações
- Av Modelagem de SistemasDocumento4 páginasAv Modelagem de SistemasWagner WittAinda não há avaliações
- Compreensão textualDocumento203 páginasCompreensão textualLuna Navarro100% (1)
- Vias de Administração FarmacoDocumento60 páginasVias de Administração FarmacoCarolina MoralesAinda não há avaliações
- Apostila CADDocumento133 páginasApostila CADProjMec100% (3)
- Trauma GenitalDocumento30 páginasTrauma GenitalWaires ZevianiAinda não há avaliações
- Be3-15 Medalha SV Mil PDFDocumento57 páginasBe3-15 Medalha SV Mil PDFGabrielaAlvesAinda não há avaliações
- Stream Via Protocolo SRTDocumento17 páginasStream Via Protocolo SRTArielly SantiagoAinda não há avaliações
- As Núpcias Celestes: A Integração das VirtudesDocumento221 páginasAs Núpcias Celestes: A Integração das Virtudesofanimenoch100% (2)
- Pombo-Gira e o resgate do sagrado femininoDocumento34 páginasPombo-Gira e o resgate do sagrado femininomoria008Ainda não há avaliações
- Catalogo Enfil Tratamento de Agua e EfluentesDocumento24 páginasCatalogo Enfil Tratamento de Agua e EfluentesMauricio LamenzaAinda não há avaliações
- Livro CC I - Calculo de Curto CircuitoDocumento76 páginasLivro CC I - Calculo de Curto CircuitoJoão Maria Staveski100% (2)
- Autoportantes VantagensDocumento3 páginasAutoportantes VantagensTecnico CoberafricaAinda não há avaliações
- Cultivo de plantas medicinais orgânicasDocumento19 páginasCultivo de plantas medicinais orgânicasDênio MonteiroAinda não há avaliações
- Convecção ForçadaDocumento17 páginasConvecção ForçadaZeca Boaventura NhachungueAinda não há avaliações
- ASP .NET - Acessando PostGreSQL em Camadas e Com Padrões de ProjetoDocumento14 páginasASP .NET - Acessando PostGreSQL em Camadas e Com Padrões de ProjetoMarceloMoreiraCunhaAinda não há avaliações
- Teste - OD - UFCD8986Documento3 páginasTeste - OD - UFCD8986Tânia BatistaAinda não há avaliações
- Juramento Da Noite EternaDocumento3 páginasJuramento Da Noite EternaBiblioteca VenusianaAinda não há avaliações
- Aula 02 Slides Os MensageirosDocumento30 páginasAula 02 Slides Os MensageirosCENPAC Centro Espírita Nympho de Paula CorreaAinda não há avaliações
- Lição 4 - Pensando em ResultadosDocumento8 páginasLição 4 - Pensando em ResultadosTiago CaiopyerAinda não há avaliações
- Questionário LP detecta defeitosDocumento5 páginasQuestionário LP detecta defeitosRaquel Pereira80% (5)
- O Beijo de Judas PDFDocumento5 páginasO Beijo de Judas PDFJuliana SteinbachAinda não há avaliações
- Lav 750 MaqDocumento2 páginasLav 750 MaqLucas MoreiraAinda não há avaliações
- Educação de Qualidade Um Desafio para MoçambiqueDocumento9 páginasEducação de Qualidade Um Desafio para MoçambiqueIgor JuriarAinda não há avaliações