Você também pode gostar
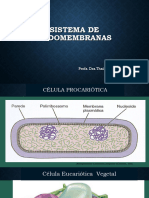
- Sistema de EndomembranasDocumento26 páginasSistema de Endomembranascarmita geronimo garciaAinda não há avaliações
- Sistema de endomembranas: componentes y funcionesDocumento9 páginasSistema de endomembranas: componentes y funcionesEdgardo Andrés Bilbao MuñozAinda não há avaliações
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDocumento7 páginasBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaAinda não há avaliações
- Complexo de Golgi e LisossomosDocumento9 páginasComplexo de Golgi e LisossomosJúlia MottaAinda não há avaliações
- Sistema de Endomembranas CelularesDocumento9 páginasSistema de Endomembranas CelularesFernanda AndradeAinda não há avaliações
- Citoplasma e OrganelasDocumento68 páginasCitoplasma e OrganelascarolsilvadornellessAinda não há avaliações
- Obtenção de Matéria PDFDocumento15 páginasObtenção de Matéria PDFLeticia RussoAinda não há avaliações
- Resumo: EndomembranasDocumento5 páginasResumo: EndomembranasBeatriz SantosAinda não há avaliações
- Sistema de endomembranas celularesDocumento4 páginasSistema de endomembranas celularesCynthia FloresAinda não há avaliações
- Resumo - GenéticaDocumento16 páginasResumo - Genéticaisabeljulia968Ainda não há avaliações
- O papel do sistema endomembranar na digestão celularDocumento7 páginasO papel do sistema endomembranar na digestão celularBoss CgAinda não há avaliações
- As Organelas Na Síntese e Degradação Das MoléculasDocumento8 páginasAs Organelas Na Síntese e Degradação Das MoléculasSamuel LemosAinda não há avaliações
- Sist - ENDOMEMBRANA 2023 - CBiomedDocumento46 páginasSist - ENDOMEMBRANA 2023 - CBiomedavenidadepirocaAinda não há avaliações
- Resumo (Citologia)Documento3 páginasResumo (Citologia)Joana LobatoAinda não há avaliações
- Organelos e transporte de proteínas nas células eucarióticasDocumento31 páginasOrganelos e transporte de proteínas nas células eucarióticasCarolina MoreiraAinda não há avaliações
- Sistema Endossômico - LisossômicoDocumento57 páginasSistema Endossômico - LisossômicoKiany de MedeirosAinda não há avaliações
- Biologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Documento65 páginasBiologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Leticia RodriguesAinda não há avaliações
- Transporte RE-GolgiDocumento25 páginasTransporte RE-GolgiAlejandro Morales GomezAinda não há avaliações
- Trabalho de Biologia CelularDocumento6 páginasTrabalho de Biologia CelularLuiz Felipe SalomãoAinda não há avaliações
- Sistema de endomembranasDocumento24 páginasSistema de endomembranasJuliana Nunes LaraAinda não há avaliações
- Biocel - PHDocumento70 páginasBiocel - PHpaulohnunesAinda não há avaliações
- Membrana BiologicaDocumento5 páginasMembrana BiologicaMaria Eduarda Padilha BaltazarAinda não há avaliações
- Roteiro SinteseDocumento4 páginasRoteiro SintesePablo MonticeliAinda não há avaliações
- Aula 3 CitologiaDocumento42 páginasAula 3 CitologiaPapelaria TeixeiraAinda não há avaliações
- Membrana Celular: Transporte e Estrutura emDocumento46 páginasMembrana Celular: Transporte e Estrutura emTatiane ValeAinda não há avaliações
- PATOLOGIA Da Celula 1Documento26 páginasPATOLOGIA Da Celula 1Sheena MendesAinda não há avaliações
- Membrana Plasmática. Digestão IntracelularDocumento27 páginasMembrana Plasmática. Digestão IntracelularBruno PereiraAinda não há avaliações
- Estudo DirigidoDocumento4 páginasEstudo DirigidoJoao Pedro Soares Amaral SousaAinda não há avaliações
- Sintese e Secreção de MacromoléculasDocumento7 páginasSintese e Secreção de MacromoléculasPablo MonticeliAinda não há avaliações
- Caio Maximus - Primeira Avaliação Do Curso de Biofísica Celular e Dos SistemasDocumento4 páginasCaio Maximus - Primeira Avaliação Do Curso de Biofísica Celular e Dos SistemasCaio MaximusAinda não há avaliações
- Aula 7 - Organelas Citoplasmáticas IIDocumento37 páginasAula 7 - Organelas Citoplasmáticas IIhome maiaAinda não há avaliações
- NST-biologia-Citologia ZeroDocumento13 páginasNST-biologia-Citologia ZeroGabriel cardosooAinda não há avaliações
- Biossíntese Do Colágeno Tipo 1Documento1 páginaBiossíntese Do Colágeno Tipo 1heguita01Ainda não há avaliações
- aula 2 biologia molecularDocumento20 páginasaula 2 biologia molecularjuuline06Ainda não há avaliações
- Via secreção biologia celularDocumento35 páginasVia secreção biologia celularKarina Fernandes ValeAinda não há avaliações
- Aula 8 - Organelas Celulares PDFDocumento42 páginasAula 8 - Organelas Celulares PDFErick SantanaAinda não há avaliações
- Biologia Geral - Aula 5 - Transporte Pela Membrana - OrganelasDocumento24 páginasBiologia Geral - Aula 5 - Transporte Pela Membrana - OrganelasJoão PauloAinda não há avaliações
- Sistema endomembranar e digestão intracelularDocumento4 páginasSistema endomembranar e digestão intracelularIurizadas0% (1)
- Membrana CelularDocumento10 páginasMembrana CelularEsmeralda MuchisseAinda não há avaliações
- Biologia - S8Documento6 páginasBiologia - S8Guilherme ColomboAinda não há avaliações
- CEDERJ Biologia Celular I - Gabarito 13 A 20Documento9 páginasCEDERJ Biologia Celular I - Gabarito 13 A 20api-3800070Ainda não há avaliações
- Resumo Biologia Celular P2Documento17 páginasResumo Biologia Celular P2Ana Beatriz Rodrigues DestefanoAinda não há avaliações
- CitologiaDocumento8 páginasCitologiaericc-13hotmail.comAinda não há avaliações
- Organização Molecular da Membrana CelularDocumento36 páginasOrganização Molecular da Membrana CelularIsabela Honorato50% (2)
- Membrana Plasmatica BioquimDocumento83 páginasMembrana Plasmatica Bioquimflavia douraoAinda não há avaliações
- Membrana celular: funções e estruturaDocumento20 páginasMembrana celular: funções e estruturaMario Maganizo JuniorAinda não há avaliações
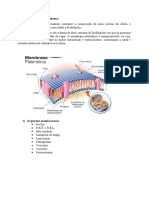
- Sistema de EndomembranasDocumento37 páginasSistema de EndomembranasRose Meire Costa BrancalhãoAinda não há avaliações
- TeoriaCitoplasma PDFDocumento9 páginasTeoriaCitoplasma PDFShofia SantosAinda não há avaliações
- 1_4931614829476577583Documento9 páginas1_4931614829476577583Saix AbelAinda não há avaliações
- BIOMEMBRANASDocumento4 páginasBIOMEMBRANASaninhayomeAinda não há avaliações
- ENDOMEMBRANASDocumento10 páginasENDOMEMBRANASYuri AlmeidaAinda não há avaliações
- Biologia Aula 2Documento8 páginasBiologia Aula 2Pierry IsseiAinda não há avaliações
- Biologia Aula 2Documento8 páginasBiologia Aula 2Kelly DomboAinda não há avaliações
- Sistema de endomembranas e organelas celularesDocumento46 páginasSistema de endomembranas e organelas celularesnoob4evermsmAinda não há avaliações
- Bio Cel Aula 3Documento5 páginasBio Cel Aula 3JoséAinda não há avaliações
- Estudo Dirigido - Reticulo Endoplasmático e LisossomosDocumento6 páginasEstudo Dirigido - Reticulo Endoplasmático e LisossomosRafael_F_AntunesAinda não há avaliações
- Estrutura Concetual Da Classificação Internacional Sobre Segurança Do DoenteDocumento145 páginasEstrutura Concetual Da Classificação Internacional Sobre Segurança Do Doentejoaopinheiro88Ainda não há avaliações
- Bases Da Radiologia ToraxDocumento35 páginasBases Da Radiologia Toraxjoaopinheiro88Ainda não há avaliações
- Anatomia Do CrânioDocumento23 páginasAnatomia Do Crâniojoaopinheiro88100% (4)
- Anatomia AbdómenDocumento21 páginasAnatomia Abdómenjoaopinheiro88Ainda não há avaliações
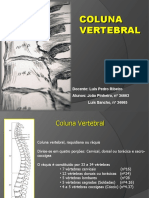
- Coluna VertebralDocumento13 páginasColuna Vertebraljoaopinheiro88100% (1)
- Texto Extra Ed Fisica 8o Ano Ens Fundamental II 17032014 1050Documento3 páginasTexto Extra Ed Fisica 8o Ano Ens Fundamental II 17032014 1050Miguel Neto71% (7)
- Catalogo EcomedDocumento46 páginasCatalogo Ecomedneo_lcAinda não há avaliações
- Sistema MuscularDocumento41 páginasSistema MuscularJoana RibeiroAinda não há avaliações
- Princípios de Podologia EquinaDocumento79 páginasPrincípios de Podologia EquinaGabriela ArmeronAinda não há avaliações
- Guia de OtoscopiaDocumento2 páginasGuia de OtoscopiaIngrid TorresAinda não há avaliações
- Anamnese completa para avaliação médicaDocumento4 páginasAnamnese completa para avaliação médicamirlenalopesAinda não há avaliações
- Mediadores Quimicos Da InflamaçãoDocumento4 páginasMediadores Quimicos Da Inflamaçãogabriela maximoAinda não há avaliações
- Ossos Membro InferiorDocumento13 páginasOssos Membro InferiorValige Pedro Valige FariaAinda não há avaliações
- Classificação Dos Tecidos, Relacionando As Suas Funções e Localização Dos Tecidos para A Formação Dos ÓrgãosDocumento26 páginasClassificação Dos Tecidos, Relacionando As Suas Funções e Localização Dos Tecidos para A Formação Dos ÓrgãosRenderson GonçalvesAinda não há avaliações
- Artrite ReumatoideDocumento6 páginasArtrite ReumatoideMaria OlíviaAinda não há avaliações
- APG 21 - PelveDocumento10 páginasAPG 21 - Pelvelarac0706100% (1)
- Hiper FLORADocumento3 páginasHiper FLORAcristressanAinda não há avaliações
- 2020 EBSOQNCPV 016 Nervos Cranianos e SentidosDocumento1 página2020 EBSOQNCPV 016 Nervos Cranianos e SentidosBenedito PurquerioAinda não há avaliações
- Semiologia Do Sistema Digestório - LivroDocumento15 páginasSemiologia Do Sistema Digestório - LivroVictoria GarciaAinda não há avaliações
- Sistema Nervoso e EndócrinoDocumento37 páginasSistema Nervoso e EndócrinoÉricaAinda não há avaliações
- Bio 2023 - Aula 01 - Introdução À CitologiaDocumento48 páginasBio 2023 - Aula 01 - Introdução À CitologiaEmily DaiannaAinda não há avaliações
- Planimetria e Terminologia Anatômica - Capítulo 3.Documento15 páginasPlanimetria e Terminologia Anatômica - Capítulo 3.Cristine Gondim MaiaAinda não há avaliações
- Sistema UrinárioDocumento12 páginasSistema UrinárioLetícia Samara Brito lsb10Ainda não há avaliações
- TD de Probabilidade 3º AnoDocumento6 páginasTD de Probabilidade 3º AnoLysiaPimentelGomesAinda não há avaliações
- Plano de Ensino Biomedicina 2 Semestre 2023 CorrigidoDocumento4 páginasPlano de Ensino Biomedicina 2 Semestre 2023 CorrigidoMONARA DE ANDRADE MARTINSAinda não há avaliações
- Cáncer de Tiroides en Nódulo Hipercaptante Por Gammagrafía. Serie de CasosDocumento6 páginasCáncer de Tiroides en Nódulo Hipercaptante Por Gammagrafía. Serie de CasosLina Marcela MoncadaAinda não há avaliações
- AD2 Botânica 1 2023 1Documento6 páginasAD2 Botânica 1 2023 1Magda GuimarãesAinda não há avaliações
- Encontro 02Documento10 páginasEncontro 02medyann09Ainda não há avaliações
- CoagulogramaDocumento39 páginasCoagulogramaverissimoliraAinda não há avaliações
- Ciências Da Pele PDFDocumento50 páginasCiências Da Pele PDFBruna Sertore Fagundes100% (1)
- APOSTILA LHCs123CarolineFelicianoMED107Documento96 páginasAPOSTILA LHCs123CarolineFelicianoMED107Gabriel MarianoAinda não há avaliações
- M.T.C. ResumoDocumento1 páginaM.T.C. Resumojailton k100% (2)
- Sistema DigestivoDocumento39 páginasSistema DigestivoDaniel José BasílioAinda não há avaliações
- Anatomia básica posições planos eixosDocumento6 páginasAnatomia básica posições planos eixosJoão EudesAinda não há avaliações
- Anatomia Do Sistema Locomotor PDFDocumento113 páginasAnatomia Do Sistema Locomotor PDFMimi Antonio100% (1)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Hackeando Darwin: Engenharia genética e o futuro da humanidadeNo EverandHackeando Darwin: Engenharia genética e o futuro da humanidadeAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Aprender mais rápido: Habilidades de estudo, memória e neurociênciaNo EverandAprender mais rápido: Habilidades de estudo, memória e neurociênciaNota: 4 de 5 estrelas4/5 (5)
- Educação ambiental: A formação do sujeito ecológicoNo EverandEducação ambiental: A formação do sujeito ecológicoAinda não há avaliações
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- Tudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNo EverandTudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNota: 4 de 5 estrelas4/5 (5)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Simetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaNo EverandSimetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaAinda não há avaliações
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Abismos Narcísicos: A Psicodinâmica do Amadurecimento e da IndividuaçãoNo EverandAbismos Narcísicos: A Psicodinâmica do Amadurecimento e da IndividuaçãoNota: 5 de 5 estrelas5/5 (2)