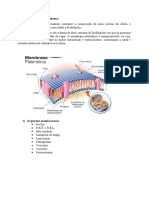
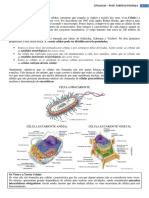
Você também pode gostar
- Tema 08 - Trafego Vesicular IDocumento35 páginasTema 08 - Trafego Vesicular IKarina Fernandes ValeAinda não há avaliações
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDocumento7 páginasBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaAinda não há avaliações
- Sistema de EndomembranasDocumento9 páginasSistema de EndomembranasFernanda AndradeAinda não há avaliações
- Via Secretória e OrganelosDocumento12 páginasVia Secretória e Organelosjoaopinheiro88Ainda não há avaliações
- Tráfego VesicularDocumento15 páginasTráfego Vesicularanonimo anonimoAinda não há avaliações
- Resmo - Tráfego Intracelular de VesículasDocumento12 páginasResmo - Tráfego Intracelular de Vesículas9vvzhn8krvAinda não há avaliações
- Compartimentos Intracelulares e TransporteDocumento8 páginasCompartimentos Intracelulares e Transportefevabax924Ainda não há avaliações
- Complexo de Golgi e LisossomosDocumento9 páginasComplexo de Golgi e LisossomosJúlia MottaAinda não há avaliações
- Roteiro SinteseDocumento4 páginasRoteiro SintesePablo MonticeliAinda não há avaliações
- Biologia P2Documento13 páginasBiologia P2Camila LimaAinda não há avaliações
- Resumo - GenéticaDocumento16 páginasResumo - Genéticaisabeljulia968Ainda não há avaliações
- Sintese e Secreção de MacromoléculasDocumento7 páginasSintese e Secreção de MacromoléculasPablo MonticeliAinda não há avaliações
- Estudo Biologia2Documento9 páginasEstudo Biologia2WQWQWEQWEQAinda não há avaliações
- CélulasDocumento5 páginasCélulasLucas NevesAinda não há avaliações
- Sistema de EndomembranaDocumento4 páginasSistema de EndomembranaCynthia FloresAinda não há avaliações
- Compartimentos Intracelulares e Endereçamento de ProteínasDocumento11 páginasCompartimentos Intracelulares e Endereçamento de ProteínasGabriel Da Silva SengesAinda não há avaliações
- Questoes BioDocumento3 páginasQuestoes Biodaniela.lima1307Ainda não há avaliações
- Biologia - Caderno de Resoluções - Apostila Volume 4 - Pré-Universitário - Biologia3 - Aula16Documento3 páginasBiologia - Caderno de Resoluções - Apostila Volume 4 - Pré-Universitário - Biologia3 - Aula16Biologia Caderno de ResoluçõesAinda não há avaliações
- Resumo de Citologia 1 PDFDocumento4 páginasResumo de Citologia 1 PDFDébora MartinsAinda não há avaliações
- Complexo GolgienseDocumento7 páginasComplexo GolgienseOtávio MayrinkAinda não há avaliações
- Resumo Biologia Celular P2Documento17 páginasResumo Biologia Celular P2Ana Beatriz Rodrigues DestefanoAinda não há avaliações
- TeoriaCitoplasma PDFDocumento9 páginasTeoriaCitoplasma PDFShofia SantosAinda não há avaliações
- Sistema de Endomembranas 2Documento9 páginasSistema de Endomembranas 2Edgardo Andrés Bilbao MuñozAinda não há avaliações
- Aula 7 - Organelas Citoplasmáticas IIDocumento37 páginasAula 7 - Organelas Citoplasmáticas IIhome maiaAinda não há avaliações
- Sistema de EndomembranarDocumento7 páginasSistema de EndomembranarBoss CgAinda não há avaliações
- FarmáciaDocumento2 páginasFarmáciaCreusa FernandesAinda não há avaliações
- CEDERJ Biologia Celular I - Gabarito 13 A 20Documento9 páginasCEDERJ Biologia Celular I - Gabarito 13 A 20api-3800070Ainda não há avaliações
- Biologia - S8Documento6 páginasBiologia - S8Guilherme ColomboAinda não há avaliações
- Ob Ten Cao de EnergiaDocumento30 páginasOb Ten Cao de EnergiaAna Margarida Lopes Alves N3 8B a3487Ainda não há avaliações
- Biologia Celular UP3 UP4Documento31 páginasBiologia Celular UP3 UP4Carolina MoreiraAinda não há avaliações
- MitocôndriasDocumento5 páginasMitocôndriasAna GrozinskiAinda não há avaliações
- Histologia - CitologiaDocumento12 páginasHistologia - CitologiaPhelipe MonteiroAinda não há avaliações
- BIOLOGIA - LisossomossDocumento2 páginasBIOLOGIA - LisossomossMariana FunghettiAinda não há avaliações
- Fichas - Organelas CitoplasmáticasDocumento6 páginasFichas - Organelas CitoplasmáticasEstefania RoblesAinda não há avaliações
- Resumo de Organelas CitoplasmáticasDocumento4 páginasResumo de Organelas CitoplasmáticasAna Laura Campos de PieriAinda não há avaliações
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Documento13 páginasModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixAinda não há avaliações
- Sistema de EndomembranasDocumento26 páginasSistema de Endomembranascarmita geronimo garciaAinda não há avaliações
- 3 - Reticulo Endoplasmático PDFDocumento15 páginas3 - Reticulo Endoplasmático PDFJoão Vitor Silva MendonçaAinda não há avaliações
- Retículo Endoplasmático RugosoDocumento1 páginaRetículo Endoplasmático RugosoPaulo ClausenAinda não há avaliações
- Membrana BiologicaDocumento5 páginasMembrana BiologicaMaria Eduarda Padilha BaltazarAinda não há avaliações
- Membrana CelularDocumento10 páginasMembrana CelularEsmeralda MuchisseAinda não há avaliações
- Bioquimica Estrutura CelularesDocumento4 páginasBioquimica Estrutura CelularesGeancarlo RibeiroAinda não há avaliações
- MI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaDocumento4 páginasMI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaEriczinAinda não há avaliações
- Estudo DirigidoDocumento4 páginasEstudo DirigidoJoao Pedro Soares Amaral SousaAinda não há avaliações
- J6 UfbddpDocumento4 páginasJ6 UfbddpNanikaGNAinda não há avaliações
- Resumo: EndomembranasDocumento5 páginasResumo: EndomembranasBeatriz SantosAinda não há avaliações
- Citologia MP e CDocumento10 páginasCitologia MP e CBird ToxicAinda não há avaliações
- Portifólio Ciclo 2Documento9 páginasPortifólio Ciclo 2Fatima aparecida de laraAinda não há avaliações
- CitologiaDocumento8 páginasCitologiaericc-13hotmail.comAinda não há avaliações
- Organelas MembranosasDocumento12 páginasOrganelas MembranosasMatheus Correa0% (1)
- As Organelas Na Síntese e Degradação Das MoléculasDocumento8 páginasAs Organelas Na Síntese e Degradação Das MoléculasSamuel LemosAinda não há avaliações
- Biossíntese Do Colágeno Tipo 1Documento1 páginaBiossíntese Do Colágeno Tipo 1heguita01Ainda não há avaliações
- 3 - MacrotransporteDocumento6 páginas3 - MacrotransporteFabiana RibeiroAinda não há avaliações
- Citologia (Rascunho)Documento33 páginasCitologia (Rascunho)marrisimoesAinda não há avaliações
- Resumão BiologiaDocumento8 páginasResumão Biologiacaroline vitoria leal silvaAinda não há avaliações
- Resumão BiologiaDocumento8 páginasResumão Biologiacaroline vitoria leal silvaAinda não há avaliações
- Especialidade PacificadorDocumento2 páginasEspecialidade PacificadorJoana LobatoAinda não há avaliações
- Feminicidio .Filosofia.Documento5 páginasFeminicidio .Filosofia.Joana LobatoAinda não há avaliações
- O Sistema CirculatórioDocumento4 páginasO Sistema CirculatórioJoana LobatoAinda não há avaliações
- Banner MastiteDocumento1 páginaBanner MastiteJoana LobatoAinda não há avaliações
- Atividade 5ano Ed. Fisica 2° Semana de SetembroDocumento1 páginaAtividade 5ano Ed. Fisica 2° Semana de SetembroJoana LobatoAinda não há avaliações
- Manual Da Conexao Com o Futuro 2022Documento19 páginasManual Da Conexao Com o Futuro 2022Joana LobatoAinda não há avaliações
- Calendario Dos Encontros Sincronos Online (Lives)Documento1 páginaCalendario Dos Encontros Sincronos Online (Lives)Joana LobatoAinda não há avaliações
- Aula 5 - ImunoglobulinasDocumento39 páginasAula 5 - ImunoglobulinasPrsjb ItaimAinda não há avaliações
- Relatorio EspermatozoidesDocumento9 páginasRelatorio EspermatozoidesJéssica RaquelAinda não há avaliações
- Massagem ClassicaDocumento38 páginasMassagem ClassicaFlorence Lainscek100% (2)
- Manobra de HeimlichDocumento34 páginasManobra de HeimlichAntenor Bispo Dos SantosAinda não há avaliações
- A Origem Das Aves: Uma Discussão ContínuaDocumento2 páginasA Origem Das Aves: Uma Discussão ContínuaMario Arthur FavrettoAinda não há avaliações
- Livro - A Força Curativa Da Respiração (Marietta Till)Documento51 páginasLivro - A Força Curativa Da Respiração (Marietta Till)maguszareth100% (7)
- Estrutura Química e Interação Molecular Farmacodinâmica Entre Salicilatos e Oxicans PDFDocumento24 páginasEstrutura Química e Interação Molecular Farmacodinâmica Entre Salicilatos e Oxicans PDFJoão CarlosAinda não há avaliações
- Cartilha Química Das Emoções 1Documento13 páginasCartilha Química Das Emoções 1lucimara de souzalima100% (4)
- MECANISMOS+DE+AÇAO+DE+HERBICIDAS+02 UnlockedDocumento52 páginasMECANISMOS+DE+AÇAO+DE+HERBICIDAS+02 UnlockedRafael FerreiraAinda não há avaliações
- Caderno de GastroDocumento123 páginasCaderno de GastroNilo Guimarães100% (1)
- Processo para Obtenção de Energia Dos Seres VivosDocumento5 páginasProcesso para Obtenção de Energia Dos Seres VivosÍtaloLuizAinda não há avaliações
- Transporte TransmembranaDocumento19 páginasTransporte TransmembranaLuciane Urman100% (2)
- Aula 02 Biologia CelularDocumento42 páginasAula 02 Biologia CelularFernanda MoraesAinda não há avaliações
- LipídiosDocumento8 páginasLipídioscamilachacurmeneguelAinda não há avaliações
- Tireoidite de HaschimotoDocumento7 páginasTireoidite de HaschimotoCamila SoaresAinda não há avaliações
- CálcioDocumento24 páginasCálcioMylena100% (1)
- Trabalho CitologiaDocumento5 páginasTrabalho CitologiaBárbara LuizaAinda não há avaliações
- Exercícios GasometriaDocumento2 páginasExercícios GasometriaJosiane VieiraAinda não há avaliações
- Desfibrilador DF 03Documento25 páginasDesfibrilador DF 03Ronaldo vieiraAinda não há avaliações
- 12t. Aula Amphibia 2014 PDFDocumento58 páginas12t. Aula Amphibia 2014 PDFYúseb MuzaarteAinda não há avaliações
- Atividade 03 Anatomia PDFDocumento2 páginasAtividade 03 Anatomia PDFSimone Da Costa CostaAinda não há avaliações
- Fisiologia VascularDocumento23 páginasFisiologia Vascularaldesblock2Ainda não há avaliações
- 03 Autismo Uma Perspectiva Sensorial 1Documento9 páginas03 Autismo Uma Perspectiva Sensorial 1Jossiane AraújoAinda não há avaliações
- Avaliação Celulas e TecidosDocumento3 páginasAvaliação Celulas e TecidosBreno MouraAinda não há avaliações
- Trabalho de BiologiaDocumento8 páginasTrabalho de Biologiadebora_wakesAinda não há avaliações
- Lista de Exercícios - 1º Estágio (2022.1)Documento51 páginasLista de Exercícios - 1º Estágio (2022.1)larissa.diasAinda não há avaliações
- Zonas de TreinamentoDocumento4 páginasZonas de TreinamentoAlexandre Rodrigo Da SilvaAinda não há avaliações
- Trabalho de Biologia - Excretor 2Documento12 páginasTrabalho de Biologia - Excretor 2World Music Group100% (1)
- Medresumos 2014 - Anatomia Humana Sistêmica 06 - Sistema CirculatórioDocumento11 páginasMedresumos 2014 - Anatomia Humana Sistêmica 06 - Sistema CirculatórioGabriel HansAinda não há avaliações
- Anatomia Topográfica Da Parede AbdominalDocumento15 páginasAnatomia Topográfica Da Parede AbdominalNoema Bastos RodriguesAinda não há avaliações