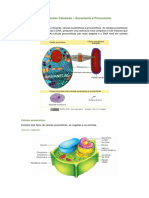
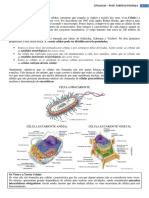
Você também pode gostar
- Organelas CitoplasmáticasDocumento5 páginasOrganelas CitoplasmáticasBrena SousaAinda não há avaliações
- UFAC 2014dDocumento16 páginasUFAC 2014dLUCES Liga Universitaria de Cirugías y EstéticaAinda não há avaliações
- Principais características dos filos de protistas Euglenida, Kinetoplastida, Ciliophora, Apicomplexa e DinoflagelladosDocumento11 páginasPrincipais características dos filos de protistas Euglenida, Kinetoplastida, Ciliophora, Apicomplexa e DinoflagelladosMinistério AliançaAinda não há avaliações
- REMIT e CicatrizaçãoDocumento4 páginasREMIT e CicatrizaçãoBarbara NevesAinda não há avaliações
- MODELO MAPA BIOLOGIA E BIOQUIMICADocumento4 páginasMODELO MAPA BIOLOGIA E BIOQUIMICAGustavo Antonio Dos SantosAinda não há avaliações
- O que é uma célulaDocumento18 páginasO que é uma célulaSilvie WacknbathAinda não há avaliações
- Organelas e estruturas citoplasmáticasDocumento7 páginasOrganelas e estruturas citoplasmáticasEduardo MottaAinda não há avaliações
- Aula 05 Sistemas de Endomembranas e OrganelasDocumento24 páginasAula 05 Sistemas de Endomembranas e OrganelasBIANCA PIFFERAinda não há avaliações
- Resumo - GenéticaDocumento16 páginasResumo - Genéticaisabeljulia968Ainda não há avaliações
- NST-biologia-Citologia ZeroDocumento13 páginasNST-biologia-Citologia ZeroGabriel cardosooAinda não há avaliações
- Biologia - Citologia - Profa KaireDocumento4 páginasBiologia - Citologia - Profa KaireKaire NardiAinda não há avaliações
- Descoberta das células e estrutura básicaDocumento33 páginasDescoberta das células e estrutura básicamarrisimoesAinda não há avaliações
- Biologia Aula 2Documento8 páginasBiologia Aula 2Pierry IsseiAinda não há avaliações
- Biologia Aula 2Documento8 páginasBiologia Aula 2Kelly DomboAinda não há avaliações
- Aula 01 - IntroduçãoDocumento43 páginasAula 01 - IntroduçãoJordeilson AmaralAinda não há avaliações
- Estudo DirigidoDocumento4 páginasEstudo DirigidoJoao Pedro Soares Amaral SousaAinda não há avaliações
- EssecitoplasmaDocumento39 páginasEssecitoplasmaAna Léa de Oliveira AraújoAinda não há avaliações
- O citoplasma das células e suas organelasDocumento52 páginasO citoplasma das células e suas organelasRaquel AcacioAinda não há avaliações
- Diferenciações da membrana plasmática e organelas citoplasmáticasDocumento4 páginasDiferenciações da membrana plasmática e organelas citoplasmáticasGeancarlo RibeiroAinda não há avaliações
- CitologiaDocumento8 páginasCitologiaericc-13hotmail.comAinda não há avaliações
- Organelas citoplasmáticas principaisDocumento5 páginasOrganelas citoplasmáticas principaisCassia RhayaneAinda não há avaliações
- Células eucariontes: características e organelasDocumento9 páginasCélulas eucariontes: características e organelasluis augusto aquatti capatoAinda não há avaliações
- Pesquisa de BiologiaDocumento4 páginasPesquisa de BiologiaEnoc FerreiraAinda não há avaliações
- O citosol e as organelas celularesDocumento22 páginasO citosol e as organelas celulareskeyse rochaAinda não há avaliações
- Organelas citoplasmáticas e suas funçõesDocumento23 páginasOrganelas citoplasmáticas e suas funçõesMariaNavesRibeiroLopesAinda não há avaliações
- Célula eucariótica e suas organelasDocumento4 páginasCélula eucariótica e suas organelasDiêgo LimaAinda não há avaliações
- Estrutura Celular ProntoDocumento4 páginasEstrutura Celular ProntoCauet CamargoAinda não há avaliações
- J6 UfbddpDocumento4 páginasJ6 UfbddpNanikaGNAinda não há avaliações
- O que é citoplasmaDocumento2 páginasO que é citoplasmaElisangela Raquel Nunes De OliveiraAinda não há avaliações
- Aula 2 CitoplasmaDocumento38 páginasAula 2 CitoplasmaJoice Caroline Lucas JuniorAinda não há avaliações
- Aps MicrobiologiaDocumento63 páginasAps MicrobiologiaAntonio FerrareziAinda não há avaliações
- Funções das organelas celularesDocumento6 páginasFunções das organelas celularessamara vitoria frias zanellato zanellatoAinda não há avaliações
- ALMENDocumento22 páginasALMENIsmael César FançaneAinda não há avaliações
- ICM - Biologia - Parte 2Documento55 páginasICM - Biologia - Parte 2Edmond TAinda não há avaliações
- Trabalho de BiologiaDocumento10 páginasTrabalho de BiologiaMaria helena De amorim ferreiraAinda não há avaliações
- Estrutura e funções das organelas celulares animaisDocumento7 páginasEstrutura e funções das organelas celulares animaisCarolina DiasAinda não há avaliações
- Aula 8 - Organelas Celulares PDFDocumento42 páginasAula 8 - Organelas Celulares PDFErick SantanaAinda não há avaliações
- Turmadefevereiro-biologia1-Tipos de Células e Membrana Plasmática-12!04!2023-Fb6326a8fa586694572784406618a143Documento17 páginasTurmadefevereiro-biologia1-Tipos de Células e Membrana Plasmática-12!04!2023-Fb6326a8fa586694572784406618a143bio-luysasantosAinda não há avaliações
- Resumo de Organelas CitoplasmáticasDocumento4 páginasResumo de Organelas CitoplasmáticasAna Laura Campos de PieriAinda não há avaliações
- Retículo EndoplasmáticoDocumento10 páginasRetículo EndoplasmáticoWillyane Samara Leandro FerreiraAinda não há avaliações
- Citologia IDocumento31 páginasCitologia IEmanuelly FajardoAinda não há avaliações
- CitologiaDocumento7 páginasCitologiaTeresa Rosaria de Souza Gonzalez EnriquezAinda não há avaliações
- Organitos e Suas Funções - JR - para PreencherDocumento2 páginasOrganitos e Suas Funções - JR - para PreencherMariana AnjosAinda não há avaliações
- Citologia 2º AnoDocumento36 páginasCitologia 2º AnoQuezia UniterAinda não há avaliações
- CITOLOGIA - TaniaDocumento7 páginasCITOLOGIA - TaniageannyyassudaAinda não há avaliações
- Citoplasma CitologiaDocumento51 páginasCitoplasma CitologiaFernando JuniorAinda não há avaliações
- Ebook Extensivo Medicina - Semana 40Documento190 páginasEbook Extensivo Medicina - Semana 40djeifnaAinda não há avaliações
- Organelas celulares e suas funçõesDocumento52 páginasOrganelas celulares e suas funçõesWallace SalvatoreAinda não há avaliações
- ResumoDocumento6 páginasResumoEulina GaldinoAinda não há avaliações
- 8ano CelulaDocumento23 páginas8ano Celulanatysmbio1173Ainda não há avaliações
- Complexo GolgienseDocumento7 páginasComplexo GolgienseOtávio MayrinkAinda não há avaliações
- Organelas CelularesDocumento9 páginasOrganelas CelularesLethícia BernardesAinda não há avaliações
- Célula Vegetal - ESALQDocumento62 páginasCélula Vegetal - ESALQAndréFerrazAinda não há avaliações
- Citoplasma e organelasDocumento273 páginasCitoplasma e organelasAluna CFP 467, Arielly, 16 pelotãoAinda não há avaliações
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDocumento7 páginasBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaAinda não há avaliações
- Citoplasma e Organelas CitoplasmáticasDocumento26 páginasCitoplasma e Organelas CitoplasmáticasBrendow FerraroAinda não há avaliações
- Aula 7 - Organelas Citoplasmáticas IIDocumento37 páginasAula 7 - Organelas Citoplasmáticas IIhome maiaAinda não há avaliações
- A diferença entre procarióticas e eucarióticasDocumento7 páginasA diferença entre procarióticas e eucarióticastrabalho ifesM27Ainda não há avaliações
- Biologia Celular: "Organelas"Documento48 páginasBiologia Celular: "Organelas"GISELE APARECIDA DA LUZ RIBAS OLIVEIRA PEREIRAAinda não há avaliações
- Microsoft Word - CitologiaDocumento3 páginasMicrosoft Word - CitologiaFelipe MagalhãesAinda não há avaliações
- Sistema de Endomembranas CelularesDocumento9 páginasSistema de Endomembranas CelularesFernanda AndradeAinda não há avaliações
- Sistema de endomembranasDocumento3 páginasSistema de endomembranasDaniela BarbosaAinda não há avaliações
- Exercicios Divisao CelularDocumento6 páginasExercicios Divisao Celularlilolok100% (1)
- 23 Metabolismo RNADocumento23 páginas23 Metabolismo RNAThayanaTabarkiewiczAinda não há avaliações
- Ae gfbf11dp Teste 1aDocumento11 páginasAe gfbf11dp Teste 1aFlavia RoxoAinda não há avaliações
- Citologia, Histologia e TecidosDocumento20 páginasCitologia, Histologia e TecidosTheDarkLight YTAinda não há avaliações
- LAKESISDocumento3 páginasLAKESISKarolyne VieiraAinda não há avaliações
- Aula 13Documento36 páginasAula 13Nuno G. PintoAinda não há avaliações
- Materia de CiênciasDocumento2 páginasMateria de Ciênciasmaria isabelaAinda não há avaliações
- Biologia Cel e MolecularDocumento62 páginasBiologia Cel e MolecularLeticia CostaAinda não há avaliações
- Sebenta de Microbiologia e ParasitologiaDocumento138 páginasSebenta de Microbiologia e Parasitologiasaramarote89100% (2)
- Pré-Universitário Oficina Do Saber Aluno (A)Documento8 páginasPré-Universitário Oficina Do Saber Aluno (A)api-518632873Ainda não há avaliações
- Teste de Biologia Ultimo 12 AnoDocumento7 páginasTeste de Biologia Ultimo 12 AnoSónia CastroAinda não há avaliações
- DNA dupla héliceDocumento8 páginasDNA dupla hélicecfernandes_7492967% (3)
- Receptores ClonaisDocumento9 páginasReceptores ClonaisOtávio GomesAinda não há avaliações
- Banco de Exercícios - Envoltórios Celulares (Recuperação)Documento8 páginasBanco de Exercícios - Envoltórios Celulares (Recuperação)Emanoel Tiago Barbosa CiriloAinda não há avaliações
- 2 - Tecido NervosoDocumento6 páginas2 - Tecido NervosoLuciano PeixotoAinda não há avaliações
- Teste - 1 - 2023 - 24 InêsDocumento10 páginasTeste - 1 - 2023 - 24 InêsjosecancelaAinda não há avaliações
- Procol La SylDocumento9 páginasProcol La SylIsteuria CristinaAinda não há avaliações
- Biologia Celular e MolecularDocumento42 páginasBiologia Celular e MolecularFernanda MoraesAinda não há avaliações
- E-Book Principais Anemias - FarmaceuticandoDocumento36 páginasE-Book Principais Anemias - FarmaceuticandoPaloma Das Candeias Dos Santos100% (1)
- Fórmulas TensorasDocumento6 páginasFórmulas TensorasDr. Luis Gonzaga AraújoAinda não há avaliações
- GlicóliseDocumento4 páginasGlicóliseGlen daAinda não há avaliações
- Estudo 02 - Atividade de estudo sobre acidose metabólicaDocumento9 páginasEstudo 02 - Atividade de estudo sobre acidose metabólicaalba_silva_54Ainda não há avaliações
- Resumo de Fisiologia - Prova Parcial 09-11-2022Documento26 páginasResumo de Fisiologia - Prova Parcial 09-11-2022Thamily OliveiraAinda não há avaliações
- Células e suas organelasDocumento4 páginasCélulas e suas organelasKARINA RODRIGUESAinda não há avaliações
- Reumatologia - 2020Documento524 páginasReumatologia - 2020nicodemosAinda não há avaliações