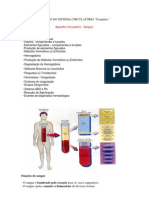
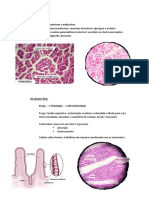
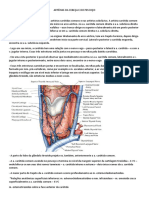
Você também pode gostar
- Assistência de Enfermagem ao Paciente com Distúrbios HematológicosDocumento8 páginasAssistência de Enfermagem ao Paciente com Distúrbios Hematológicosingrid.naruto.fbAinda não há avaliações
- Fisiologia Do SangueDocumento22 páginasFisiologia Do SangueVanessa ArrudaAinda não há avaliações
- Livro HematologiaDocumento36 páginasLivro HematologiaNathalia Guimarães NogueiraAinda não há avaliações
- Aula 07 - Tecido SanguíneoDocumento8 páginasAula 07 - Tecido SanguíneoGabriella ASAinda não há avaliações
- HematoloiaDocumento6 páginasHematoloiarafael souzaAinda não há avaliações
- O SangueDocumento13 páginasO SangueNathan GuilhermeAinda não há avaliações
- Medula Óssea e HematopoeseDocumento54 páginasMedula Óssea e HematopoeseRafa Arshavin100% (2)
- Células Do Sangue e HemocitopoeseDocumento10 páginasCélulas Do Sangue e HemocitopoeseGuilherme NascimentoAinda não há avaliações
- Fisiologia da circulação sanguíneaDocumento44 páginasFisiologia da circulação sanguíneaGleidsonRodriguesAinda não há avaliações
- AULA1Documento37 páginasAULA1Faizal abacar SaideAinda não há avaliações
- Introdução Á Hematologia IDocumento25 páginasIntrodução Á Hematologia IMaria ClaraAinda não há avaliações
- Histologia Do Sangue - Células Do Sangue - Resumos MedicinaDocumento17 páginasHistologia Do Sangue - Células Do Sangue - Resumos MedicinaAdriana CravoAinda não há avaliações
- Resumo - Sistema Circulatório e Sistema RespiratórioDocumento17 páginasResumo - Sistema Circulatório e Sistema RespiratórioThayane MoreiraAinda não há avaliações
- Capítulo 13 Robbins PatologiaDocumento7 páginasCapítulo 13 Robbins PatologiaCarolina StrongerAinda não há avaliações
- RESUMODocumento13 páginasRESUMOKim BandeAinda não há avaliações
- TEMA 1 OkDocumento27 páginasTEMA 1 OkDaniela LimaAinda não há avaliações
- Processo de hematopoiese e formação de células sanguíneasDocumento5 páginasProcesso de hematopoiese e formação de células sanguíneasLucasAinda não há avaliações
- Módulo II - Unid. 05 HematologiaDocumento44 páginasMódulo II - Unid. 05 HematologiaMarcelaAinda não há avaliações
- Apostila 1 Hemato para impressão - 2 (2019)Documento4 páginasApostila 1 Hemato para impressão - 2 (2019)LEILA TEIXEIRAAinda não há avaliações
- Células Do Sangue PDFDocumento91 páginasCélulas Do Sangue PDFLaura QueirozAinda não há avaliações
- Composição Do SangueDocumento9 páginasComposição Do SangueMA TE USAinda não há avaliações
- Hematopoiese: formação e desenvolvimento das células sanguíneasDocumento23 páginasHematopoiese: formação e desenvolvimento das células sanguíneasAna Maria SilvaAinda não há avaliações
- Funções do sangue e elementos figuradosDocumento28 páginasFunções do sangue e elementos figuradosOilson SartoriAinda não há avaliações
- Hematologia Basica Parte 3 - HematopoeseDocumento16 páginasHematologia Basica Parte 3 - HematopoeseP2P3ESTATISTICA CIPTUR100% (1)
- Origem e Formação Dos LeucócitosDocumento3 páginasOrigem e Formação Dos LeucócitosAline PaulinoAinda não há avaliações
- HematopoieseDocumento3 páginasHematopoieseSara Veiga AndradeAinda não há avaliações
- Fisiologia Do Sistema CardiovascularDocumento32 páginasFisiologia Do Sistema CardiovascularNúbia SantosAinda não há avaliações
- Produção dos elementos sanguíneosDocumento99 páginasProdução dos elementos sanguíneosRafaela VilelaAinda não há avaliações
- Hematologia IDocumento100 páginasHematologia ItkmyjftnmxAinda não há avaliações
- Apostila Interpretação de Exames Laboratoriais INAESPDocumento38 páginasApostila Interpretação de Exames Laboratoriais INAESPCarlos Leopoldo S. FerreiraAinda não há avaliações
- Apostila Tecido Sanguà Neo - CNMDocumento9 páginasApostila Tecido Sanguà Neo - CNMDébora GuedesAinda não há avaliações
- Histologia do sangue emDocumento16 páginasHistologia do sangue emLarissa Barboza CardosoAinda não há avaliações
- Histologia do tecido sanguíneo e suas célulasDocumento39 páginasHistologia do tecido sanguíneo e suas célulasSabrina Rabelo100% (1)
- RESUMO M5 ImunologiaDocumento54 páginasRESUMO M5 Imunologiayezafig4001Ainda não há avaliações
- Fagocitose, Pinocitose e LeucócitosDocumento4 páginasFagocitose, Pinocitose e LeucócitosMatheus EPAinda não há avaliações
- Trabalho Citologia-5Documento47 páginasTrabalho Citologia-5kynhoAinda não há avaliações
- Aula 11classeDocumento11 páginasAula 11classenelsonaugusto496Ainda não há avaliações
- Auxiliar de laboratório hematologiaDocumento38 páginasAuxiliar de laboratório hematologiaMirlane Andrade costaAinda não há avaliações
- Resumo Hematologia BásicaDocumento9 páginasResumo Hematologia BásicaBiouma MadeiraAinda não há avaliações
- Sistema circulatório: transporte de oxigênio e nutrientes pelo sangueDocumento15 páginasSistema circulatório: transporte de oxigênio e nutrientes pelo sangueApolônio Sobrinho100% (1)
- Apostila Histologia 40Documento28 páginasApostila Histologia 40tailanelucianoAinda não há avaliações
- HematopoieseDocumento6 páginasHematopoieseYaslénio PopAinda não há avaliações
- Bioquímica AplicadaDocumento19 páginasBioquímica AplicadaÉvelly ValérioAinda não há avaliações
- Sistema CirculatórioDocumento69 páginasSistema Circulatórioxicana100% (3)
- 11 3 Sistema Circulato Rio Teoria Questo Es Orientadas PDFDocumento12 páginas11 3 Sistema Circulato Rio Teoria Questo Es Orientadas PDFMaria Luiza Silva FariasAinda não há avaliações
- Trabalho Biologia (2)Documento5 páginasTrabalho Biologia (2)Biah AmorimAinda não há avaliações
- AlfaCon Sistema CirculatórioDocumento10 páginasAlfaCon Sistema Circulatórioelaine santosAinda não há avaliações
- Hematologia - Formação do sangue e Eritrocitos (1)Documento5 páginasHematologia - Formação do sangue e Eritrocitos (1)LEILA TEIXEIRAAinda não há avaliações
- Fisiologia Aplicada Exercicio Sobre SangueDocumento5 páginasFisiologia Aplicada Exercicio Sobre SangueAnderleyBHAinda não há avaliações
- Sangue v1.0Documento29 páginasSangue v1.0Sílvia NevesAinda não há avaliações
- Conteúdo 3 HematopoieseDocumento6 páginasConteúdo 3 HematopoieseAdriana SilvaAinda não há avaliações
- Sistema Hematopoiético - Formação e Destruição das Células do SangueDocumento5 páginasSistema Hematopoiético - Formação e Destruição das Células do SangueCarlosGomesAPAinda não há avaliações
- Resumo Cremoso HematologiaDocumento3 páginasResumo Cremoso HematologiaMarcos Antonio D'Queiroz JuniorAinda não há avaliações
- 10342728032012Histologia_Basica_Aula_9Documento12 páginas10342728032012Histologia_Basica_Aula_9hoteldonaanarececaoAinda não há avaliações
- O Sangue: Funções e Componentes emDocumento2 páginasO Sangue: Funções e Componentes emDayanne PinheiroAinda não há avaliações
- Conceitos HematologiaDocumento14 páginasConceitos HematologiarodrigocamargovetAinda não há avaliações
- As principais leucemias: tipos, causas e sintomasDocumento34 páginasAs principais leucemias: tipos, causas e sintomasGustavo BittencourtAinda não há avaliações
- Apostila 2ºbim 8ºanoDocumento6 páginasApostila 2ºbim 8ºanoRolnan100% (1)
- Exame físico da cabeça e pescoçoDocumento64 páginasExame físico da cabeça e pescoçoJulyany RochaAinda não há avaliações
- Suseme EndocrinologiaDocumento61 páginasSuseme EndocrinologiaAgatha Uchoa BatistaAinda não há avaliações
- Procedimentos de exames e terapias especiais cobertosDocumento24 páginasProcedimentos de exames e terapias especiais cobertosJESSICA POLIZERAinda não há avaliações
- 57 180 PBDocumento290 páginas57 180 PBHanna Lícios Vieira AlvesAinda não há avaliações
- Lugol Por Que Suplementar?Documento2 páginasLugol Por Que Suplementar?linelumber3Ainda não há avaliações
- FuncionalDocumento174 páginasFuncionalSamuel FernandesAinda não há avaliações
- Respostas hormonais ao exercícioDocumento13 páginasRespostas hormonais ao exercícioWenes Pereira X Amanda EstevaoAinda não há avaliações
- Algoritmo MGFamiliar - Nódulos TiroideusDocumento2 páginasAlgoritmo MGFamiliar - Nódulos Tiroideuslittlegirl586Ainda não há avaliações
- Portfólio - Anatomia HumanaDocumento14 páginasPortfólio - Anatomia Humanalucasgabriel7162Ainda não há avaliações
- Chakras 2Documento22 páginasChakras 2Vitor LanaroAinda não há avaliações
- SUPLEMENTAÇÃO DE IODO Com Solução de LugolDocumento18 páginasSUPLEMENTAÇÃO DE IODO Com Solução de LugolWeslley Alves100% (1)
- Nódulos TireoidianosDocumento5 páginasNódulos Tireoidianosana paula de castro costaAinda não há avaliações
- Exame Da TireoideDocumento17 páginasExame Da Tireoidebarbara SA Carvalho ReisAinda não há avaliações
- Testes de Triagem NeonatalDocumento4 páginasTestes de Triagem NeonatalSuelen LimaAinda não há avaliações
- Pequenos 2Documento37 páginasPequenos 2ingrid mirandaAinda não há avaliações
- O que é a tireoideDocumento4 páginasO que é a tireoideCaio FerreiraAinda não há avaliações
- Padrões UltrassonográficosDocumento23 páginasPadrões UltrassonográficosFernanda Palermo Hassan100% (2)
- Solvana Usui New Terra Nível 1Documento20 páginasSolvana Usui New Terra Nível 1VanesssAinda não há avaliações
- Aula Sistema Genital Feminino e EndócrinoDocumento78 páginasAula Sistema Genital Feminino e EndócrinoKarinny GonçalvesAinda não há avaliações
- APR2 2º TRIM - 8º Ano CFB - GABARITO PDFDocumento6 páginasAPR2 2º TRIM - 8º Ano CFB - GABARITO PDFAntonio Correia100% (1)
- Sistemas endócrinos e desenvolvimento glandularDocumento3 páginasSistemas endócrinos e desenvolvimento glandularArthur CastroAinda não há avaliações
- Doenças NutricionaisDocumento70 páginasDoenças NutricionaisLogain ExcluiAinda não há avaliações
- Slides Doenas Endocrinas Doencas TireoidianasDocumento130 páginasSlides Doenas Endocrinas Doencas TireoidianasSergio MeloAinda não há avaliações
- Lista 04 - Estrutura AtômicaDocumento6 páginasLista 04 - Estrutura AtômicaNatália Fernanda Schuh100% (1)
- Sistema endócrino e hormônios produzidos pelas glândulasDocumento6 páginasSistema endócrino e hormônios produzidos pelas glândulasraquel luzAinda não há avaliações
- Ajustes fisiológicos da gravidezDocumento10 páginasAjustes fisiológicos da gravidezHermenegildo chitumbaAinda não há avaliações
- Processo Seletivo 2023-2: EDITAL DIRPS #09/2023 Tipo 2 1 FASE - 02/07/2023Documento44 páginasProcesso Seletivo 2023-2: EDITAL DIRPS #09/2023 Tipo 2 1 FASE - 02/07/2023Otávio AlvesAinda não há avaliações
- Resumo Prova Prática 1 - Lâminas Citologia 2016Documento4 páginasResumo Prova Prática 1 - Lâminas Citologia 2016lidiaAinda não há avaliações
- Doença de GravesDocumento77 páginasDoença de GravesYav KAinda não há avaliações
- Artérias da cabeça e do pescoçoDocumento7 páginasArtérias da cabeça e do pescoçoHenriqueAinda não há avaliações