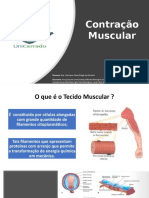
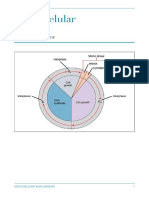
Você também pode gostar
- Transporte Vesicular Resumo AlbertDocumento5 páginasTransporte Vesicular Resumo AlbertMacklecio OliveiraAinda não há avaliações
- Estudo Dirigido de Fisiologia Vegetal E Biotecnologia - 01: D) Potencial HídricoDocumento6 páginasEstudo Dirigido de Fisiologia Vegetal E Biotecnologia - 01: D) Potencial Hídricobruno souza gonçalvesAinda não há avaliações
- Comunicação e Sinalização CelularDocumento14 páginasComunicação e Sinalização CelularGonçalo MachadoAinda não há avaliações
- WEB Ar at A05 Apostila Inter.Documento20 páginasWEB Ar at A05 Apostila Inter.diane50% (2)
- Questionario BiofísicaDocumento3 páginasQuestionario BiofísicaBruna MelloAinda não há avaliações
- Arquitetura MolecularDocumento24 páginasArquitetura MolecularMarcone MachadoAinda não há avaliações
- 2G11 Bases Puricas e PirimidicasDocumento8 páginas2G11 Bases Puricas e PirimidicasjulyannegoesAinda não há avaliações
- PEROXISSOMOSDocumento12 páginasPEROXISSOMOSelizangelacarAinda não há avaliações
- Aula Pratica 1 - Teste Viabilidade MTTDocumento2 páginasAula Pratica 1 - Teste Viabilidade MTTCarla PereiraAinda não há avaliações
- Resumos de BioenergéticaDocumento22 páginasResumos de BioenergéticaHugo NicolauAinda não há avaliações
- Aula 2 - Organelas CelularesDocumento52 páginasAula 2 - Organelas CelularesSibele FernandesAinda não há avaliações
- Cadeia RespiratóriaDocumento60 páginasCadeia RespiratóriaAlline RodriguesAinda não há avaliações
- Sebenta de Biologia Celular e Molecular FCNAUP 2019Documento98 páginasSebenta de Biologia Celular e Molecular FCNAUP 2019D CostaAinda não há avaliações
- Cadeia Transportadora de Elétrons e Fosforilação Oxidativa PDFDocumento48 páginasCadeia Transportadora de Elétrons e Fosforilação Oxidativa PDFDiesSyca Masson MassonzinhaAinda não há avaliações
- 2 Prova de BioquímicaDocumento6 páginas2 Prova de BioquímicaMary FreitasAinda não há avaliações
- Testes de Conhecimento PRINCÍPIOS DE QUÍMICA BIOLÓGICADocumento58 páginasTestes de Conhecimento PRINCÍPIOS DE QUÍMICA BIOLÓGICAMarcos MedeirosAinda não há avaliações
- Tecido NervosoDocumento14 páginasTecido NervosohaugustwAinda não há avaliações
- Membrana Plasmática - Função - Permeabilidade Da MembranaDocumento14 páginasMembrana Plasmática - Função - Permeabilidade Da MembranaHigor BarrosAinda não há avaliações
- Atividade Fase Clara e Escura FotossínteseMarcoDocumento2 páginasAtividade Fase Clara e Escura FotossínteseMarcoMarco AntônioAinda não há avaliações
- Aminoácidos e Proteínas PDFDocumento5 páginasAminoácidos e Proteínas PDFAndreAinda não há avaliações
- Replicação, Transcrição e TraduçãoDocumento37 páginasReplicação, Transcrição e TraduçãoNatalia Maria MilhoratiAinda não há avaliações
- Trabalho Contração Muscular FinalDocumento37 páginasTrabalho Contração Muscular FinalNaillyne GalvãoAinda não há avaliações
- Relatorio Bioquimica EnzimasDocumento13 páginasRelatorio Bioquimica EnzimasMax Ribeiro50% (2)
- Citoplasma e Organelas Citoplasmáticas - SlidesDocumento47 páginasCitoplasma e Organelas Citoplasmáticas - SlidesAnna BeatryzAinda não há avaliações
- Estudo DirigidoDocumento5 páginasEstudo DirigidoLaianaCarmanini100% (2)
- Aula2 Purificacao e Determinacao EstruturasDocumento69 páginasAula2 Purificacao e Determinacao EstruturasVale EuropeuAinda não há avaliações
- Cadeia RespiratóriaDocumento2 páginasCadeia RespiratóriaRaquel Cavalcanti Figueiredo100% (1)
- 3 - TranscriçãoEProcessamentoRNA PDFDocumento17 páginas3 - TranscriçãoEProcessamentoRNA PDFYago LuisAinda não há avaliações
- Aula1 - DNA e CromossomosDocumento34 páginasAula1 - DNA e CromossomosTwo Crazy React100% (1)
- Os Anfíbios - BiologiaDocumento5 páginasOs Anfíbios - BiologiaMaxvpllAinda não há avaliações
- Biologia MolecularDocumento15 páginasBiologia MolecularDaniela SayuriAinda não há avaliações
- Resumo de CnidariaDocumento9 páginasResumo de CnidariaHyago Marcello OchôaAinda não há avaliações
- Polonio PDFDocumento2 páginasPolonio PDFwozikAinda não há avaliações
- Hemocitopoese - ResumoDocumento7 páginasHemocitopoese - ResumoRichardson Oliveira50% (2)
- Estudo Dirigido - FisiologiaDocumento6 páginasEstudo Dirigido - FisiologiaFelipe AssunçãoAinda não há avaliações
- Capítulo 3 - Estrutura e Funcionamento Da CélulaDocumento48 páginasCapítulo 3 - Estrutura e Funcionamento Da CélulaMegan Katherine Melling100% (2)
- Química PPT - CASD - Moleculas - PolaridadeDocumento11 páginasQuímica PPT - CASD - Moleculas - PolaridadeQuímica PPT100% (4)
- Estudo Dirigido Matriz Extracelular e Sinalizac807a771o CelularDocumento3 páginasEstudo Dirigido Matriz Extracelular e Sinalizac807a771o CelularRafaelAinda não há avaliações
- Potencial Elétrico ResumoDocumento2 páginasPotencial Elétrico ResumoIasmim MaiaraAinda não há avaliações
- Sinalizacao CelularDocumento19 páginasSinalizacao Celulartorquato4976Ainda não há avaliações
- Gabarito de Biologia Celular - Prova 01Documento3 páginasGabarito de Biologia Celular - Prova 01Thaís LuzAinda não há avaliações
- Biofísica MolecularDocumento6 páginasBiofísica MolecularSimon PjAinda não há avaliações
- AULA 5 EstereoquímicaDocumento36 páginasAULA 5 EstereoquímicawillnerothAinda não há avaliações
- Cronograma EnemDocumento6 páginasCronograma EnemJessyca MeloAinda não há avaliações
- Celulas SanguineasDocumento9 páginasCelulas Sanguineaselizangelacar100% (1)
- Tecido ConjuntivoDocumento6 páginasTecido ConjuntivoVicttoria MeloAinda não há avaliações
- Citologia Revestimento Externo Das CélulasDocumento3 páginasCitologia Revestimento Externo Das CélulasThiago RibeiroAinda não há avaliações
- Biosíntese Do ColágenoDocumento3 páginasBiosíntese Do ColágenoGlendse PinheiroAinda não há avaliações
- Contaminação de Solo (Bioventilação)Documento9 páginasContaminação de Solo (Bioventilação)dennyele0% (1)
- Tecido Muscular 2015Documento57 páginasTecido Muscular 2015Webert Martins MeloAinda não há avaliações
- PPT22 - Obtenção de EnergiaDocumento23 páginasPPT22 - Obtenção de EnergiaMartim FranciscoAinda não há avaliações
- Exercicos AminoácidosDocumento10 páginasExercicos AminoácidospscacaoAinda não há avaliações
- Estudo Dirigido CitoDocumento4 páginasEstudo Dirigido CitoTiago SantosAinda não há avaliações
- Núcleo, Diferenciação e Morte CelularDocumento69 páginasNúcleo, Diferenciação e Morte CelularAmanda CristinaAinda não há avaliações
- GlicóliseDocumento3 páginasGlicóliseTay100% (1)
- Aula 2 - Biofísica Da ÁguaDocumento29 páginasAula 2 - Biofísica Da ÁguaKelly_Simi_9916100% (1)
- Teoria AtomicaDocumento15 páginasTeoria AtomicaJanaína MatosAinda não há avaliações
- Eletroforese em GelDocumento3 páginasEletroforese em GelKananda Lima Barreto100% (1)
- Prof Alcides SN2 SN1Documento21 páginasProf Alcides SN2 SN1Marcelo Augusto Pereira JanuárioAinda não há avaliações
- Biologia Molecular 2Documento8 páginasBiologia Molecular 2Gabriel Da Silva SengesAinda não há avaliações
- Cópia de Resumo Membranas Celulares - Aula 1Documento19 páginasCópia de Resumo Membranas Celulares - Aula 1Gabriel Da Silva SengesAinda não há avaliações
- Cópia de Resumo Membranas Biológicas - Aula 1Documento3 páginasCópia de Resumo Membranas Biológicas - Aula 1Gabriel Da Silva SengesAinda não há avaliações
- Ciclo CelularDocumento10 páginasCiclo CelularGabriel Da Silva SengesAinda não há avaliações
- BIOMORFO - Histologia Resumo 02Documento16 páginasBIOMORFO - Histologia Resumo 02Gabriel Da Silva SengesAinda não há avaliações
- Compartimentos Intracelulares e Endereçamento de ProteínasDocumento11 páginasCompartimentos Intracelulares e Endereçamento de ProteínasGabriel Da Silva SengesAinda não há avaliações
- BIOMORFO - Histologia Resumo 02Documento16 páginasBIOMORFO - Histologia Resumo 02Gabriel Da Silva SengesAinda não há avaliações
- Biologia Molecular 2Documento8 páginasBiologia Molecular 2Gabriel Da Silva SengesAinda não há avaliações
- Ciclo CelularDocumento10 páginasCiclo CelularGabriel Da Silva SengesAinda não há avaliações
- Biologia Molecular 2Documento8 páginasBiologia Molecular 2Gabriel Da Silva SengesAinda não há avaliações
- Cópia de Resumo Membranas Biológicas - Aula 1Documento3 páginasCópia de Resumo Membranas Biológicas - Aula 1Gabriel Da Silva SengesAinda não há avaliações
- Reumato Roteiro 12 11 2019Documento6 páginasReumato Roteiro 12 11 2019pedro albertoAinda não há avaliações
- Ciclo CelularDocumento10 páginasCiclo CelularGabriel Da Silva SengesAinda não há avaliações
- Exame Do JoelhoDocumento18 páginasExame Do JoelhoJulio CezarAinda não há avaliações
- Exame Do JoelhoDocumento18 páginasExame Do JoelhoJulio CezarAinda não há avaliações
- AvaljoelhoDocumento15 páginasAvaljoelhopeurenatinha100% (1)
- Portifolio 3 Pronto Biologia Primeiro SemestreDocumento6 páginasPortifolio 3 Pronto Biologia Primeiro SemestreDênis CechinelAinda não há avaliações
- Linfadenite Caseosa em Caprinos e OvinosDocumento13 páginasLinfadenite Caseosa em Caprinos e OvinosAline KoslinskiAinda não há avaliações
- Odis10 Teste4 CorrecaoDocumento2 páginasOdis10 Teste4 CorrecaoAna MoutinhoAinda não há avaliações
- Obtenção de Matéria Pelos Seres HeterotróficosDocumento16 páginasObtenção de Matéria Pelos Seres HeterotróficosDulce SantosAinda não há avaliações
- Questões para A Prova Da DanieleDocumento7 páginasQuestões para A Prova Da DanieleDanieleTeixeiraCostaAinda não há avaliações
- Estrutura CelularDocumento31 páginasEstrutura CelularTales bruno Goncalves costaAinda não há avaliações
- Manual Osmose Reversa Smart - REV04Documento25 páginasManual Osmose Reversa Smart - REV04Adailton Marinho100% (1)
- Ficha Formativa - TransportesDocumento3 páginasFicha Formativa - TransportesElisaAinda não há avaliações
- Relato de Experiência KahootDocumento12 páginasRelato de Experiência KahootDAYANE CANDIDO DA SILVAAinda não há avaliações
- Trabalho de Antibioticos-CARIMODocumento18 páginasTrabalho de Antibioticos-CARIMOCarimo gorges ArmandoAinda não há avaliações
- Lista de Exerccios Sobre Potenciais de Repouso e de Ao Da Clula ImagensDocumento2 páginasLista de Exerccios Sobre Potenciais de Repouso e de Ao Da Clula ImagensThiago de AndradeAinda não há avaliações
- Uniceluridade MulticelularidadeDocumento34 páginasUniceluridade MulticelularidadeinesAinda não há avaliações
- Farmacocinética - Absorção e DistribuiçãoDocumento12 páginasFarmacocinética - Absorção e Distribuiçãoauriculoterapia.wesAinda não há avaliações
- 1º Bimestre - Biologia - 3aDocumento97 páginas1º Bimestre - Biologia - 3aanakaroliny.zx021Ainda não há avaliações
- Atividade Lipideos e Carboidratos Com RespostaDocumento2 páginasAtividade Lipideos e Carboidratos Com RespostaFrancine MendesAinda não há avaliações
- Ciclo Do FósforoDocumento6 páginasCiclo Do Fósforohillbill2009100% (2)
- Simulado BIOQUÍMICADocumento14 páginasSimulado BIOQUÍMICAMatheusAinda não há avaliações
- Estruturas Internas Da Célula Bacteriana Aula 2Documento22 páginasEstruturas Internas Da Célula Bacteriana Aula 2Daniela Ricardo MuradasAinda não há avaliações
- Texto de Apoio de Biologia Evolutiva Sobre Evidencias EvolutivasDocumento10 páginasTexto de Apoio de Biologia Evolutiva Sobre Evidencias EvolutivasDavid JoaoAinda não há avaliações
- Lista Revisão Biologia 1° E.MDocumento9 páginasLista Revisão Biologia 1° E.Mrecomenda booksAinda não há avaliações
- Captura de Tela 2023-06-26 À(s) 5.09.05 PMDocumento11 páginasCaptura de Tela 2023-06-26 À(s) 5.09.05 PMLucieli MarquesAinda não há avaliações
- Landmark4216 en PTDocumento13 páginasLandmark4216 en PTPedro Henrique SantosAinda não há avaliações
- Anatomia e Fisiologia para Psicólogos - Carlos Alberto PastoreDocumento52 páginasAnatomia e Fisiologia para Psicólogos - Carlos Alberto PastorebarbaraAinda não há avaliações
- A Célula Como Unidade Da VidaDocumento23 páginasA Célula Como Unidade Da VidaAlberto Jose MbanzeAinda não há avaliações
- CélulasDocumento4 páginasCélulasizinhasoaresAinda não há avaliações
- Guia IlustradoDocumento43 páginasGuia IlustradoLorenaAinda não há avaliações
- Apostila de Biologia ENEM 1Documento11 páginasApostila de Biologia ENEM 1Ana030982Ainda não há avaliações
- Golan 1 Interacoes Farmaco ReceptorDocumento16 páginasGolan 1 Interacoes Farmaco ReceptorDaniel Sant AnaAinda não há avaliações
- Prova BM320Documento4 páginasProva BM320Yanara OliveiraAinda não há avaliações
- Neurofisiologia 3Documento42 páginasNeurofisiologia 3Karine de FreitasAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Hackeando Darwin: Engenharia genética e o futuro da humanidadeNo EverandHackeando Darwin: Engenharia genética e o futuro da humanidadeAinda não há avaliações
- Aprender mais rápido: Habilidades de estudo, memória e neurociênciaNo EverandAprender mais rápido: Habilidades de estudo, memória e neurociênciaNota: 4 de 5 estrelas4/5 (5)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- Tudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNo EverandTudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNota: 4 de 5 estrelas4/5 (5)
- Dieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNo EverandDieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNota: 5 de 5 estrelas5/5 (4)
- Uma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNo EverandUma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNota: 4.5 de 5 estrelas4.5/5 (9)
- O Licenciamento Ambiental no Brasil na PráticaNo EverandO Licenciamento Ambiental no Brasil na PráticaNota: 1 de 5 estrelas1/5 (2)