CAPITULO 8
Existem duss condigdes fundamentals para a vida, Uma deltas &
que 0 organismo vivo deve ser capar de se auto-replicar (topico
que serd visto na Parte IV deste livro). A segunda ¢ que 0 orga-
nismo deve ser capaz de catalisar reagOes guimicas eficiente €
seletivamente. A importancia fundamental ca catdlise pode sur-
preender alguns estudantes iniciantes em bioquimica, mas € fa-
dil ilustra-la, Como descrito no Capitulo 1, 08 sistemas vivos
tilizam energia do meio ambiente, Osseres humanos, por exem-
plo, consomem quantidades substanciais de sacarose — 0 agti-
‘ar comumente usado — como uma espécie de combustivel, na
forma de alimentos arucarados, bebidas,ou como acticar. A con-
versio da sacarose em CO; € HO na presenca de oxigénio é um
processo altamente exergimnico, que libera energia livre que pode
ser uiilizada para o ato de pensar, mover, sentir e ver. Entretan-
to, um saco de agticar pode ser armazenado durante varios anos,
sem ser transformado em CO, e HO, Embora esse processo qui-
smico seja termodinamicamente favordvel, ele é muito lento! Con-
tudo, quando a sacarose é consumida por uma pessoa (ou qual-
‘quer outro organismo}, ela libera sua energia quimica em se-
gundos. A diferenca é a catdlise. Sem a catdlise, as reacdes
‘quimicus necessirias para sustentar a vida no ocorrem em uma
«scala de tempo util.
Voltaremos, agora, nossa atengio para os catalisadores das
1eagbesque ovorrem nossistemas biolégicos: as enzimas, es mais
notdveis ¢ altamente especializadas proteinas. As enzimas apre-
sentam uma eficiéncia catalitica extraordindria, em geral muito
‘maior que a dos catalisadores sintéticos ou inorganicos. Flas ém
um alto grau de especificidade por seus substratos, aceleram as
reagdes quimicas de uma maneira formidaével ¢ funcionam em
solgdes aquosas sob condicdes muito suaves de temperatura €
PH, Poucos catalisadores nao-biolégicos apresentam tais pro-
Priedades.
{As enzimas so fundamentais para qualquer processo bio-
quimico. Agindo em seqGéncias onganizadas, elas catalisam as
‘entenas de reagOes sucessivas pelas quais as moléculas nutrien-
tes sio degradadas, a energia quimica ¢ conservada e transfor-
mada ¢ as macromoléculas biolégicas so sinteiizedas a partir
de moléculas precursors simples. Devido 3 a¢io das enzimas
reguladoras, as vias metabdlicas so altamente coordenadas de
forma a produzir uma atuacso harmoniosa das muitas diferen-
tes atividades necessirias para manter a vida.
© estudo das enzimas tem uma grande importincia prética,
Em algumas doengas, especialmente nas desordens genéticas
herdadas, pode ocorrer uma deficitneia ou mesmo a auséncia
‘otal de uma ou mais enzimas, Outras condigoes anormais tam-
bém podem ser causadas pela excessiva atividade de uma emi
‘ma, Medidas da atividade de enzimas no plasma sangiineo, ri
trécitos ou amostras de tecido sto importantes no diagnéstico
de virias doencas. Muitas drogas exercem seu efeito bioldgico
por meio de interagoes com as enzimas. As enzimas tornaram-
139
imas
se importantes ferramentas priticas nio apenas na medicina
mas também na industria quimica, no processamento de ali-
mentos e na agricultura.
Iniciaremos este capitulo com a descrigéo das propriedades
das enzimas e dos principios findamentais de seu poder catali-
tio. Em seguida, faremos uma introducio a cinética enzimati
ca, uma disciplina que fornece a maior parte dos fundamentos
para qualquer discussio sobre enzimas. Exemplos especificns de
mecanismos enzimiticos serao entdo mostrades, ilustrando os
Principios introduzidos anteriormente. Finalmente, discutire-
mos as enzimas reguladoras.
Uma Introducao as Enzimas
A maior parte da historia da bioquimica ¢ a historia da pesquise
sobre enzimas. A catdlise biolégica foi inicialmente reconhecida
descrita no final de 1700, em estudos sobre a digestao da carne
por secregdes do estéimago. Em 1800 as pesquisas continuaram
com @ estudo da conversio do amido em agticares simples pela
saliva € por virios extratos vegetais. Em 1850, Louis Pasteur con-
cluiu que a fermentagio do asticar em ileool pela levedura é ca
talisada por “fermentos” Ele postulou que esses fermentos eram
insepardveis da estrutura das células vivas de levedura, uma hi-
puitese chamada vitalismo, que prevaleceu por muitos anos. Em.
1897, Eduard Buchner descobriu que os extratos de levedura
podiam fermentar o agicar até dleool, provando que a fermen.
tagao era promovida por moléculas que continuavam funcio-
nando mesmo quando removidas das células. Frederick W. Kuh
ine denominou tais moléculas de enzimas. A medida que as no-
‘ees do vitalismo da vida foram refutadas, oisolamento de novas
cenzimas e 2 investigagio das suas propriedades propiciaram 0
progresso da citncia da bioquimica.
Eduard Buchner
(1850-1917)
isolamento ea cristalizagao da ureese, em 1926, por James
Sumner, propiciaram uum significante avanco nos estudos ini-
ciais acerce das enzimas, Sumner demonstrou que os cristais de
urease consistiam inteiramente de proteina ¢ postulou que to-
das as enzimas sio proteinas. Devido falta de outros exemplos,
essa premissa permaneceu controvertida por algum tempo. So-
190
James Sumner
11887-1955) 18S. Haldane
(1892-1968)
mente em 1930, aps John Northrop ¢ Moses Kunitz terem cris-
talizado a pepsina, 2 tripsina e outras enzimas digestivas ¢ con-
dluido que elas também eram proteinas, é que a conclusio de
Sumner foi completamente aceita. Durante esse period, |.B.S.
Haldane escreveu um tratado intitulado “Enzimas”. Embora a
natureza molecular das enzimas mo estivesse completamente
lucidade, Haldane fez a extraordindria sugestao de que as inte-
racies fracas que se estabelecem entre a enzima eo seu substra-
to poderiam ser usadas para distorcer a molécula do substrato ¢
«atalisar 2 reagao, Esse conhecimento representa 0 cemne da com-
preensio atual da catalise enzimitica
No final do século XX, a pesquisa com enzimas que catali-
sam as reacies do metabolismo celular foi intensa. Esse fato le-
voua purificagio de milharesde enzimas, aclucidagioda estrutu-
ra molecular e do mecanismo quimico da acio de centenas delas
€a.uma compreensio geral de como as enzimas funcionam.
A maioria das enzimas so proteinas
Com excegio de um pequeno grupo de moléculas de RNA que
apresentam propriedades cataliticas (Capitulo 26), todas as en-
zimas sio proteinas. A sua atividade catalitica depende da inte-
gridade da sua conformasio protéica nativa. Se uma enzima €
desnaturada ou dissociada em subunidades, a atividade cataliti-
«a geralmente é destruida. Se uma enzima ¢ quebrada em scus
aminodcidos constituintes, @ sua atividade catalitica € sempre
destraida. Assim, as estraturas protéicas primatia, secundétia,
tercidria ¢ quaterndria das enzimas sao essenciais para a sua ati-
vidade catalitica.
‘As enzimas, como outras proteinas, ttm pesos moleculares
{que variam de cerca de 12,000 até mais de 1 milhao. Algumas
enzimas ndo requerem nenhum outro grupo quimico, além de
seus tesiduos de aminodcidos, Outras requerem um componente
po
C-£,C—5, C_Oe CN pormeia de
reacBes de condersatao accoladas 8
‘queora co ATP
"a raona dis encimas catasa a transferenca de eletons, ators oa grupos
funcionals. Dasse modo, elas s2oclass‘icedes de acordo com umm nimero de
fd. e atribu ndo-se umn nome de acordo como ipo reaydo de trarsteré-
,c9 oruno deadore do grupo acepter,
mero na Comissao de Enzimas (mimeo B.C.) 2.7.1.1. 0 pri-
meiro digito (2) denota o nome da classe (transferase); 0 se-
sgundo digito (7), a subclasse (fasfotransferase): 0 terceiro digi-
to (1), uma fosfotransferase que apresenta um grupe hidroxila
aceptor de fosfato; ¢ 0 quarto digito (1) indica que a D-glicase ¢
aceptor do grupo fosfato. Pars muitas enzimas, um nome tri-
val pode ser usado, como por exemple no caso da hexoquinase.
Uma lista completa e a descricao das milhares de enzimas
conhecidas esté além do objetivo deste livro, Este capitulo é des-
tinado principalmente aos principios e as propriedades comuns
todas as enzimas.
Como as Enzimas Funcionam
Acatélise enzimética das reacbes ¢ essencial para os sistemas vi-
9s. Sob condigdes biolégicas relevantes, as reagbes nao catali-
sadis tender a ser lentas — muites das motéculas biol6gicas
sio bastante est4veis no ambiente aquoso, de pH neutro e tem
prratura moderads, do interior das células. Além disso, muitas
reacdes bioquimicas comuns envolvem eventos quimicos que
sio desfavoriveis ou improvaveis no ambiente celular tais como
1 formacao de intermedisrios carregados ou a colisto de duas
(1 mais moléculas com a orientacio precisa necessaria para que
‘corra a reagdo, Sem catélise, as reagbes necessérias para digerir
5 alimentos, enviar sinais através dos nervos ou contrair um
iiculo simplesmente nao ocorrem com uma velocidade ‘itil
‘Uma enzima contorna esses problemas fornecendo um am-
biente especifico onde uma dada reacio é energeticamente mais
favordvel. A caracterfstica que distingue uma reagio catalisada
enzimaticemente € a de ela ocorser no interior dos limites de
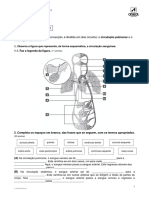
tums cavidade na enaima chamada sitio ativo (Fig. §-1).A molé-
«ak que se liga ao sitio ativo e sofre a agao da enzirva ¢ chamada
substrato. A superficie do centro ativo é contornada com resi-
duos dc aminoacids cujos grupos substituintes se ligam ao subs-
{rato e catalisam a sua transformacao. O complexo enzima-subs-
trato, cuj existéncia foi inicialmente proposta por Adolphe
Wurtz em 1880, ¢ fundamental para a agao das enzimas. Ele €
também o ponto de partida para os tratamentos matemiticos
que definem © comportamento cinético das reag6es catalisadas
por enzimas, bem como para as descrig&es tedricas dos meca-
nismos enzimitices.
191
Figura 0-1 ~ Ligagdo de uma molécula de substrate no sitio ative
dde ume enzima. A enzima quimatripsna esta ligada ao substrate rros
‘vadoem vermelno, Alguns aminoacicos essericals do sitia avo $20 ros.
‘rados como manchas vermelhas ra superficie de enzima
As enzimas afetam a velocidade mas
ngo 0 equilibrio quimico das reaces
Uma reacao enzimética simples pode ser escrita
4S ESP SE
wy
onde E, § ¢ P representam, a enzima, 0 substrato e 0 produto,
sespectivamente. ES EP sao complexos transientes da enzima
com o substrato e a produto.
Para compreender « catilise enzimética, precisamos enten-
det primeiro a importante distincdo entre equilibrio da reacao
{(discutido no Capitulo 4) e velocidade da reagio. A fungio de
um catalisador € aumentar a velocidade de uma reagio. Os cata-
lisadores nio afetam 0 equilibrio da reacéo. Qualquer reacio do
tipo SP pode ser descrita por um diagrama de coordenadas
da reacao (Fig. 8-2), uma representagdo das mudancas de ener-
gia durante a reagio. Conforme explicado nos Capitulos 1 e3,a
éenergia nos sistemas biologicos ¢ descrita em termos de energia
livre, G. No diagrama de coordenadas, a energia livre do sistema
€ representada em fungio do progresso da reagdo (coordenada
da reac), O ponto de pattida, tanto para a reagao de ida como
para a reagio reversa, é chamado estado fundamental « repre-
senta acontribuigaa de uma molécula tpica(S ou P) para aener-
gia livre do sistema, em um dado conjunto de condigdes, Para
Energia live, G
Coordenedas de reagio
Figura 8-2 ~ Dlagrama de coordenadas de reacso para uma reagso
‘quimica, A energia lite do sisterna @ representada em func0 do pro-
aresso da reacio S==*P. Um diaarama desta natureza descreve as mu-
ddangas de energia curante a tesco. e 0 eixa horizontal icoordenadas ca
reagio) reflete as miudarcas quimicas sucessvas (por exerrplo, quabra e
formagto de figacées) 3 medida que S & corvertido ern PAs eneraias de
ativagao para as (2ag0es 5 -» Pe P—» Sesto indkcadas por AG". AG™ ea
vaniagao total da energia lvre padrao para 3 reacao SP
192
descrover as varingbes de energia livre das reagdes, os quimicos
definem um conjunto de condigdes-padao {temperatura 298K;
pressio parcial de cada gis 1 atm ou 101,3kPa; concentragao de
cada soluro IM) ¢ expressam a variagdo da energia livre para
esse sistema reagente como AG", a variagdo da energia livre pa-
drao. Como os sistemas biolégicos geralmente envelvers con-
centragies de H" muito diferentes de IM, os biogaimicas defi-
nem z variagdo da energia livre padrao bioquimice, AG", come
sendo a variagio da energia livre padrio em pH 7.0, que sers
utilizada neste livio, Uma definigdo mais completa de AG” ¢
dada no Capitulo 14,
‘© equilbbrio entre S e ? reflete a diferenga em energia livre
dos seus estados fundamentais. No exemplo mostrado na Figu-
+1 8-2, 2 energia livre do estado fundamental de P é menot que a
de 5, assim AG" & negative para a reagio Sr P ¢ 0 equilibrio
favorece a formagao de P. A posigao e a direcao do equilibrio nae
so afetadas por nenhum catalisador.
‘Umequilfbrio favoravel nao significa que aconversioS— P
‘ocorra com uma velocidade mensurivel. A velocidade da tea-
‘gio depende de um parametro completamente diferente. Existe
luma barreira energética entre S e P que representa a energia
necessiria para o alinhamento dos grupos quimicos reagentes,
formagio de cargas tronsientes instaveis, rearranjos de ligagoes
outras transformacdes necesséries para que 3 reagao ocorra
fem uma das diregSes. Isso é ilustrado pela “colina” de energia
pas Figuras 8-2 ¢ 8-3. Para softer a reagdo, as moléculas deve
superar essa barreira ¢, portanto, precisam ser excitadas até um
nivel de energia maior. No topo da colina de energia esté um
ponte ro qual a passagem para o estado de 5 ou P¢ igualmente
provével (é sempre um caminho “morro abaixo” em cada senti-
€o}, Esse € 0 chamado estado de transigio. O estado de transi
io nao é uma espécie quimica que apresenta uma estabilidade
significativa e no deve ser confundido com um intermediario
«a reasio (tal como ES ou EP), Fle é aperas um momento mo-
lecular efemero no qual eventos como quebra de ligacées, for-
‘magi de ligagdes.e desenvolvimento de cargas ocorrem em um
ponto preciso no qual a decomposicao para formar o substrato
(016 produto é igualmente provavel. A diferenca entre os niveis
de energia do estado fundamenial ¢ do estado de transigio €
chamada energia de ativagdo (AG'). A velocidade de uma rea-
fo reflete essa energia de ativasao, isto é, ume energia de ativa-
ae alta corcesponde a uma reagzo lenta, As velocidades des re-
agdes podem ser aumentadas pela elevarao da temperatura, que
aumenta 0 niimero de moléculas com energia suficiente para
superar essa barreira de energia, Alternativamente, 2 energia de
ativacao pode ser diminuida pela adicao de um catalisador (Fig
8-2). Os catalisadores aumentan a velocidade das reagdes dimi
nuindo a energin de ativacio,
As enzimas nio so excegio & regea de que os catalisadores
nao afetam o equilibrio da reaczo. As setas bidirecionais na Equa-
‘8-1 deixam claro este ponto: qualquer enzima que catalisa a
reagdo § ~> P também catalisa a reagao P -> S.O papel das enzi
mas ¢ acelerara interconversio de Se P. A enzima nao ¢ gasta no
processo € o ponto de equilfbrio nao é afetado. Entretanto, a
reagdo atinge 0 equilibrio muito mais rapidamente quando a
entima apropriada esta presente, uma vez. que a velocidade da
reagio é aumentada.
Esse principio geral pode ser ilustrado considerando-se a
conversio da sacarose e oxigénio em CO, € H.0:
Cutl0,, + 120, == 12005 + 11,0
sia conversdo, que ovorre por meio de uma sétie de reagdes
isoladas, tem um AG” elevado e negativo, ¢, no equilibrio, a
quantidade de sacarose é despreaivel, A sacarose é um composto,
Coordenadas de veayao
Figura 6-3 Dlagrama de coordenadas de teagéo comparando uma,
reagio catallsada enzimaticamente com uma ndo-catalisada, Ne
feacao 5» P05 intermediarios ES EP assumem valores mnimos na
‘curva de progresso da reacao catalisada enzimaticamente. Os termos
Ginn € AGin Covrespondem as eneiaias de ativacto das reacoes na0-
calalseda @ catalsada, respectivamente. A energia de atwacto para 0
procesto global ¢ menor cuando a enzima catalsa a rexgio,
estével porque. harreira de energia de ativagao que deve ser supe-
rada para que ela reaja com o oxigtnio é muito grande. Ela pode
ser armazenada em um recipiente contendo O, indefinidamente,
sem reagir. Entretanto, nas células, 2 sacarose é rapidamente
transformada em CO; ¢ 1,0 por meio de uma série de reajoes
catalisadas por enzimas, Essas enzimas nao apenas aceleram as
reagOes, mas as organizam e as controlam de tal forma que a
‘maior parte da energia liberada é conservada em outras formas
quimicas, ficando assim disponivel paraa célula executar outras
tarefas. O caminho reacional pelo qual a sacarose (¢ outros acu-
cares) é degradada 6 a via priméria de liberagio de energia (Capt-
tulos 15 19), €as enzimas dessa via permitem que asequencia de
reagbes oenrra em uma escala de tempo biologicamente itil,
Na pritica, qualquer reagdo pode ter varios passos (etapas)
envolvendo a formagio e 0 constimo de espécies quimicas tran-
sientes, chamades de intermedisrios da reagio*. Quando a res-
20.5 P € cataliseda por uma enzima, 0s complexos ES ¢ EP
sio os intermediirios (Eg, 8-1). les ocupam os vales no diagra-
ma de coordenadas da reegao (Fig, 8-3). Quando ocorrem vérios
passos em uma reagio, a velocidace total € determinada pelo
passo (ou pessos) com a maior energia de ativagio. Esse passo é
chamado “passo limitante da yelocidade’, Para os casos simples,
6 paseo limitante da velocidade € © ponto de mais alta energia
no diagrama para as interconversoes de S e P.INa pratica, © passo
limitante da velocidade pode variar com as condigdes da reacio.
Para muitas enzimas, varios passos tém energia de ativacao si-
ilar. o que significa que todos eles sto parcialmente limitantes
da velocidace.
Como cescrite no Capitulo 1, as energias de ativagio sio
barreiras energéticas para as reagdes quimicas. Essas barreiras
sao cruciais para a existencia da propria vida. A estabilidade de
‘uma molécula aumenta com o aumento da altura de sua barr
ra ce ativagao. Sem tais barreiras energeticas, as macromoléca-
las complexas poderiam reverter espontaneamente para as for-
‘mas moleculares mais simples, e es estruturas complexas alta-
* Observe que os termos “passes” e“intermediiios” neste capitulo se r=
ferem as espécies quimicas que ecorrem no caminho reacional de uma
simples rea;ao catalisada enzimaticamente. No contexio das vias meta-
bolicas que envelver virins enzimas (Parte IIT deste livr), esses terms
sao usados de uma maneita diferente, Uma reagso eneitatica gelmet
6 considerada como um “passo” em uma via metabélica, ¢ 0 produto de
‘uma reagao enzimética (que ¢ substrato para a pr6xima ensima na via)
considerado coma um “intermedisrio"
mente ordenadas, bem como 0s processos metabélicos das célu-
las, poderiam nao existir. As encimas evoluiram para diminuit
seletivamente 2s énergias de ativag2o de reaches necessérias A
sobrevivencia das células.
A velocidade ¢ 0 equilibrio das reagdes tém
definigoes termodinamicamente precisas
Ocquilibrio de uma reagao esta intrinsecamente ligado ao AG®,
enquanto a velocidade da reagao esta ligada 20 AG". Uma intro-
dugio bisica a essas relagies termodinamicas & 0 préiximo pas-
so na compreeasio de como as enzimas trabalham,
Um ecuilibrio como $= P € descrito por uma constante
de equillbrio, K., ou simplesmente K (Capftulo 4). Sob condi-
goes-padrao, empregadas para comparar processos bioqut
os, uma constante de equilibrio ¢ expressa por Ki, (ou K")
teh
‘s)
i (82)
Da termodinamica, a relagdo entre Kt, e AG” pode ser descrita
pela expressao:
AGP = RT In Ky (8.3)
onde 2 éa constante dos gases, 8,315]/mol -K, e Téa tempera-
{ura absoluta, 298K (25°C). A Equagao 8-3 sera desenvolvida €
discutida com maiores detalhes no Capitulo 14. O ponto im-
portante aqui é que a constante de equilibrio csté dirctamente
relacionada com a varia¢do global da energia livre padrao da
eagle (Tabela 8-4). Um grande valor negativo para AG" reflete
tum equilibrio de reagao fayoravel mas, como ja foi salientado,
isso nio significa que a reacio ocorrers com uma velocidade
considerdvel,
Tabela 8.4 ~ A relacdo entre Kig ¢ AG” (veja Eq. 8-3)
quena fragio de segundo, Se a velocidade da reagio depende da
concentragio de dois compostos diferentes, ou se duas moléculas
do mesmo composto reagem entre si, a reacao & de segunda or-
dem e k € uma constante de velocidade de segunda ordem, com
umidade M's, Nesse caso, a equacdo da velocidade torna-se:
¥=AS115o 65)
[A partir da teorie do estado de transigao, pode-se derivar uma
expressdo que relacionaa magnitude da constantede velocidade
com a energia de ativagior
ke KE pacer
wo
onde k éa constante de Boltzmann ¢ h &a constante de Planck.
Oponto importante a salientar aqui é que arelagao entre acons-
tante de velocidad, f, ¢ @ energia de ativagio, AG', é inversa €
exponencial, Em outras palavras, essa é a base para a afirmagdo
de que uma energia de stivag3o menor significa uma velocidade
de reacao maior e vice-versa.
Agora vamos mudar 0 assunto de o que as enzimas fizem,
para como elas 0 fazem.
Alguns principios explicam 0 poder catalitico
ea especificidade das enzimas
As enzimas sAo catalisadores extraordinarios. Os aumentos da
velocidade provocados pelas enzimas variam entre $ ¢ 17 ordens
de magnitude (‘Tabela 8-5). As enzimas também sao muito es-
pecificas, discriminando entre substratos com estruturas muito,
similares. Como esses aumentos, enormes ¢ dltamente seletivos,
na velocidade das reagdes podem ser explicados? De onde vema
‘energia que diminui dramaticamente a energia de ativagao para
reagdes especificas?
Tabela 8-5 - Alguns valores do aumento da velocidade produzido
Ke ‘AG (kd/mol) por enzimas.
iy Ea Cidlofilina 10°
108 : .
ee a Anidiase carbonica 10
a tat Tose fostato komerase 108
107 114 Caboxipentidese A 10”
tot a Fosfoglicomutase 10?
1 00
uciniLCOA transferase 108
it ba Succi CoA trans 0"
a0 ana Urease 10"
0 12.1 rotidina monofastato descarboxlase 10”
‘A velocidade de qualquer reacio é determinads pela con-
centragio do reagente (ou dos reagentes) ¢ pela constante de
velocidade, geralmente representada pelo stmbolo k, Para a rea-
fo unimolecular $ -> P,a velocidade da reagao ¥, que represen.
taaquantidade deS que reage por unidade de tempo, ¢ expressa
pela equagio da velocidade:
v=uis] (ea)
Nesta reagio, a velocidade depende apenas da concentragio de S.
Hlaé chamada de reagao de primeira ordem. O fator ké uma cons-
tante de proporcionalidade que reflete a probabilidade de a rea-
«fo ocorrer em um dado conjunto de condicoes (pH, temperatu-
racte,). Aqui, k 6uma constante de velocidade de primeira ordem
sua unidade ¢ o reciproco do tempo, por exemplo 5 '. Se uma
reacio de primeira ordem tem uma constante k de velocidade de
0,038", isso pode ser interpretado (qualitativamente) que 39 do
S disponivel seri convertida a P em 1s. Uma reagdo com uma
comstante de velocidade de 2.000s'" estar terminada em uma pe:
‘A resposta para essas questdes apresenta duas partes distin-
tas, mas interligadas. A primeira diz respeito aos rearranjos das
ligacoes covalentes durante a reacdo enzimatice. Reagdes quimi-
cas dos mais variados tipos ocorrem entre o substrato ¢ os gru-
_pos funcionais da enzima (cadeias laterais de aminodcidos espe-
cificos, fons metilicos e coenzimas). Os grupos funcionais cats-
liticos das enzimas podem formar uma ligayao covalente
transitéria com um substrato e ativé-lo paraa reacio ou, entdo,
algum grupo pode ser transferido transientemente do substrato
pata um grupo da enzima. Em muitos casos, essas reagdes s0-
‘mente ocorrem no centroative éa enzime. Elas diminuem a ener-
‘gia de ativacio (¢, portanto, aceleram a reagao), propiciando um.
caminho reacional akternativo de energia mais baixa.
‘A segunda parte da resposta diz respeito as interagdes nao
covalenies entre a enzima e a substrato. A maior parte da ener-
gia necessiria para diminuir a energia de ativaydo € devivada
de interacbes fracas, ndo-covalentes, entre 0 substrato e @ enzi-
ma. O fator que de fato distingue as enzimas da maioria dos
193
194
catalisadores nao-enziméticos ¢ a formagie de um complexo
ES especifico. A interacdo entre o substrato e a enzima nesse
complexo é mediada pelas mesmas forgas que estabilizam a
trutura protéica, incluindo as pontes de hidrogénio, inters-
gies hidrofébicas e idnicas (Capitulo 6). A formasao de caca
interagdo fraca no complexo ES € acompanhada por uma pe-
quena liberacao de energia livre que garante o grav de estabili-
dade para a interagao, A energia derivada da interagao encime-
substrato & chamada de energia de ligagio, AGy. O seu signifi-
cado vai além de uma simples estabilizagao da interagio
enzima-substrato. A emergia de ligagao é a maior fone de ene
aia Fire usada pelas encinias para diminuir a energia de ativa-
(io das reagées
Doisprincfpios fundameniais einter-retacionadosfornecem
tama explicagio geral para fato deas enzimas atilizarem acner
gia de ligagao nao-covalente.
1. A maior parte do poder catalitieo das enzimas é derivada, em
ultima instancia, da energia live liberada na formagio das mail-
tiplasligagdes fracas einteragées entre a enzima eo seu substra
to. Essa energia de ligagio contribui tanto para a especificidade
comp para a catalise
2,.As interagdes fracas sio otimizadas no estado de transigao da
reacio. Os centros ativos das enzimas so complementares no
40s rexpectivos substratos per si, mas acs estados de transicao
pelos quais os substratos passam a medida que so transforme-
dos em produtos durante as reagdes enzimatices
Esses aspectos sio eritices para a compreensao da agae das enzi-
mas ¢ agora eles se tornardo 0 foca da nossa atencao,
As interagdes fracas entre a enzima e 0 substrato
so otimizadas no estado de transicio
Como uma enzima utiliza a energia de ligagao para diminuir a
energie de ativago da reagdo? Embara a formacao do comple-
x0 FS nao seja a explicagio adequada, algumes das primeiras
consideragdes dos mecanismos enzimaticos comegaram com
essa premissa. Os estudos sobre a especificidade das enzimas
deseavolvidos por Emil Fischer levarami-no a propor, em 1894,
Figura
{que as encimas eram estruturalmente complementares aos seus
substratos, de tal forma que se ajustariam fal qual “fechadura ¢
chave” (Fig. 8-4),
Essa premissa elegante de que a interagio especifica (exclu:
siva) entre duas moléculas biolégicas é mediada por superficie:
meleculares com formas complementares influenciou fortemente
« desenvalvimento da bioguimica e tais interagdes permanecert
no cerne de muitos processes bioquimicos. Entrelanto, a hipé-
tese “fechactura e chave" pode gerar confusao quando aplicada i
catiliseenzimities, Uma enzima totalmente complementara sev
substrato seria urna enzima muito pouco eficiente,
Considere uma reagio imaginaria, a quebra de um bastic
ce metal magnetizado. A reagio ndo-catalisada esti mostrade
na Figurad-5a. Vamos analisar duasenzimasimagindrias —duas
“hastonases” — que catalisam esta reigio e ambas empregando
forgas megnéticas como modelo para a energia de ligagao em-
pregada pelas enzimasreais, Yamos considerarinicialmente uma
enzima perfeitamente complementar ao substrato (Fig. 8-56).
© centro ative dessa “bastemase” ¢ uma cavidade delimitada por
‘magnetos. Para reagir (quebrar),o bastio precisa atingir o esta
do de transigio da reagdo, mas ele se ajusta muito firmemente
ab sitio ativo que nao pode se dobrar. Isso porque o dobramen-
to do bastao climinaria parte das interagdcs magnéticas entre
dle € a enzima, Esse tipo de enzima impede « reagio, uma vez
‘que, na realidade, estabiliza o substrate, Num diagrama de co-
‘ordenadas da reagao (Fig. 8-3b), esse tipo de complexo ES cor-
responderia a um pogo de energia do qual o substrato difiil-
mente escaparia, Esse tipo de enzima seria inatil.
‘A nogao moderna de catalise entimatica, proposta inicial-
mente por Haldane em 1930, foi elaborada por Linus Pauling,
com 1946:a fim de catelisar reagdes, uma enzima deve ser com-
plementar a0 estade de transigao. Isso significa que as intera-
5 timas (por meio de ligagdes fracas) entre o substrato ¢ a
enzima ocorrem apenas no estado de transigio. A Figura 8-5¢
mostra como uma enzima desse tipo pode trabalhar, O bastio
dde metal liga-se a enzima, mas algumas poucas interagbes mag-
néticas so usadas para formar o complexo ES. © substrato lic
gaco precisa ainda receber um aumento de energie livre para
Formas complementares de um substrato e seu sitio ativo em uma enzima A enzima dildrofolato edutase é mostrade
om o seu substrato NADP" (vermelno) n3o igado (3 esquerd e igado (a diel), Também € wsivel um autro substato lgado & encima. 0
‘etraidrofclato iamarelo). © NADP" lga-se a uma cavidade que € complementar &sua forma e propriedades inicas. Na tealicace, a comple
mmentaridade entre 2 pr
‘ina @ 9 igante (neste caso, o substrato| raremente & parfeita, como jd wste no Copttuls 7. A mteracdo de uma
Protens com um ligante frequentemante envolve mudangas na confermacao de Uma ou ambas ae meléculas, um pracesto dencmriade
juste nduzido, Esse falta de uma complementaridade perfelta entiea encima e o substrata (nao eviderciada nesia figura) e importante pare
2 catalise enzimatice
{a} Sem enzima
ee — 4% _. i
== g ®y 3
Substrate Estado de transigto Produtos 2
(Chastio de metal) (basta dobrado) (hastao partido) =
(>) Enzima complementar a0 substrato
2
fmas €
2
pee 3
=
a
(©) Enzima complementar ao estado de transigio-
4
Figura 8-5 - Uma enzima imagindria ("bastonase") projetada para catalisar a quebra de um bastio de metal. (a) Para ser
‘quebrado, primeiramente 0 bastic deve sar dabrado (o estado de transicio). Em ambos 0s exermpios de “bastonase”, as interacbes
‘nagnéticas fazer 0 papel das nteracoes por ligagbes fracas entre enzima e substrato. (b) Ua “bastonase” que apresenta uma
‘ayidade magretizada com estrutura Corrplemerta’ d estrutura do bastBo (0 substato) estabilza esse substrata, O dobramento do
bastao ¢ impedido pela aracdo magnetica entre a bestao e a “bestonase™. (© Uma enzima complementar a0 estado de transiao
da reagao ajuda 2 desestabilzar 0 bastao, coninbundo para a calalse da rearao. A energia de ligaca0 das interagoes magreticas
compensa o aumento de energiaInre necesséria para dobrar 0 basta. Os dagramas de coordenadas da reacao (8 direta) mostram
a consequtncias energeticas da complementaridade ao substrato versus 2 complementaridade 20 estado de transx0. O termo
AGu representa a diferenca entre 25 energas dos estados de transigao da reaczo néo-catalisada e da catazada € provém das
interaches magnecas entre © bastio e 2 “bastonste”. Quando a enzime & complementar ao substato (b), 0 complexo ES 6 mai
‘estvele tem menos energi Inte no estado fundamental que o proprio substrata. O resultado ¢ um aurmento na energia de atnacBo.
‘Coordenudas de reas
195
stingir © estado de transicio. Assim, o cumento em energia li-
‘re necessirio para dobrar o bastao © quebrar parcialmente a
sua conformaclo ¢ compensado ou “pago” pelas interagdes
rmagnéticas (enengia de ligagdo) que se formam entre aenzimae
‘o substrato no estado de transicdo. Muitas dessas interacoes
envolvem partes do bastdo que esto longe do ponto de rompi-
mento. Assim serido, is imteragdes entre a"bastonase” as regioes
‘nio-reativas do bastio fornecem parte da energia necessiria para
‘orompimento do bastio. Esse "pagamento de energia” é tradlu-
ido em uma menor energia de ativacio real e uma maior velo-
Gidade de reagio.
‘As erzimas reais trabalham segundo um principio anlogo.
Algamas interagdes fracas sio formadas no complexo ES, mas a
totelidade das interagSes fraces que se estabelecem entre 0 subs-
trato ¢ a enzima ¢ estabelecida apenas quando o substrato atin=
g@ 0 estado de transipao. A encrgia livre (energia de ligngio) li-
berada pela formagao dessas interagoes supre parcialmente a
‘energia requetida para se atingir 0 topo de calina de energia. ©
somatério da cnergia de ligacao desfavordvel (positiva), MG €
favordvel (negativa), AG, resulta em uma energia de ativagio
liguida menor (Fig, 8-6). Mesmo para a enzima,o estado de tran-
sido ndo € uma espécie estavel, mas um breve instante que o
‘Coordenadas de rexgio
Figura 8-6-0 papel da energla de ligacio na catélise, Para dimiruir
a energa de ativacio de uma reagio, © sistena precisa adquinr uma
quanidade de energie equvvlente a diminugdo de AG" maior parte
essa eneiga provem da energa de Iqacao |AGs) e @ dsponbitzede
pela formacéo de intetagbes nao-cavalentes fracas enire o substato e 3
enzima no estado de transcé0. O signficaca de AGy & analogo a0 de
AGy na Figura 8-5.
196
substrato gasta para atingir o topo da barreira de energia. A rea-
‘s20 catalisada enzimaticamente € muito mais répida que © pro-
‘cesso nao-catalisado, porque a barreira é muito menor. O prin-
cipio importante ¢ que as interagies de ligacdes fracas entre a
encima ¢ 0 substrato fornecent a maior parte da forga que dirige a
catdlise enzimética. Os grupos no substrato que sio envolvides
nessas interagdes fracas podem estar a uma certa distancia das
ligagoes que s40 quebradas ou mudadas. As interacOes fracas que
se formam apenas no estado de transicio sio as que contribuem.
fandamentalmente para a catilise.
‘A necessidade de miltiplas interacdes fracas para dirigit a
catilise ¢ uma das razbex por que as enzimas (e algumas coenzi-
‘mas) sto moléculas tao grandes. A enzima deve fornecer 0s gru-
pos funcionais para a formacao de interagdes iénicas, pontes de
hidrogénio ¢ outras interagdes, bem como posicionar precisa-
‘mente esses grupos de tal forma que a energia de ligacao seja
otimizada no estado de transigao.
Aenergia de ligacao contribui para
a especificidade da reagéo e a catélise
E possivel demonstrar quantitativamente que a energia de liga-
‘40 6 responsivel pela enorme aceleragio da velocidade das rea-
{G0es catalisadas pelas enzimas? Sim. Como um ponto de refe-
réncia, a Equacao 8-6 permite calcular que AG" deve diminuir
aproximadamente 3,7kJ/mol para acelerar cerca dedez vezes uma
reagio de primeira ordem, sob as condicdes comumente encon-
tradas nas células. A energia disponivel devido 3 formagio de
‘uma Unica interagao fraca geralmente € da ordem de 4 2 30ki/
mol. A energia total disponivel, devido a formagio de um certo
niimero-dessas interagics, provavelmente € suficiente para di-
‘minuir a energia de ativagao de cerca de 60 a 100k)/mol, reque-
rida para explicar os grandes aumentos na velocidade cbserva-
dos para muitas enzimas.
‘A mesma energia de ligacao que fornece a energia para a
catélise também € responsével pela especificidede, a habilidade
da enzima em discriminar entre o substrato e uma outra molé-
cula competidora. Conceitualmente, « especificidade é facil de
ser diferenciada da catalise, mas essa distingao € muito mais di-
ficil de ser feita experimentalmente, porque a catalise ¢ a especi~
ficidade sio originérias do mesmo fendmeno. Se © centro ativo
dde uma enzima tem grupos funcionais arranjados de forma oti-
smizada para formar uma variedade de interagies fracas com um
dado substrato mo estado de transigio, a enzima nao sera capaz
de interagir do mesmo modo com qualquer outra molécula. Por
‘caemplo, seo substrato tem um grupo hidroxila que forma uma
ponte de hidrogénio especifica com um residuo de écido gluti-
‘mico na enzima, qualquer molécula que ndo possuir aquele gra
po htdroxila especifico sera, de maneira geral, um substrato mais
pobre para a mesma enzima. Além disso, qualquer molécula com
tum grupo funcional extra, para o qual a enzima néo apresenta
uma cavidade ou um sitio de ligacdo, muito provavelmente sera
‘excluida da enzima. Geralmente, a especificidade & derivads da
formagao de mitiplas interagbes fracas entre 2 enzimae a mo-
leécula de seu substrato especifico.
(Os prinetpios geraisjé descritos podem ser ilustrados por
luma grande variedade de mecanismos cataliticos conhecidos,
Esses mecanismos nao sao mutuamente exclusivos e uma dada
‘enzima pode incorporar virios deles em seu mecanismo de ago,
Geralmente ¢ dificil quantificar a contribuigdo de um dado me-
canismo catalitico para velocidade e/ou especificidade de uma
determinada rea¢ao catalisada enzimaticamente,
‘A energia de ligagio € a forga impulsionadora dominante
«em varios mecanismos ¢ la pode ser 0 principal, se ndo 0 tinico,
contribuinte para a catdlise, Vamos considerar 0 que € necessi
rio acontecer para que a reasio ocorra. Os fatores fisicos e ter-
modinamicos importantes que cantribuem para 0 AG", a bar-
reira energética, incliem: (1) a mudanga na entropia, na forma
‘de movimento de liberdade de duas moléculas em solugio; (2) a
‘camada de solvatacdo de moléculas de igua ligadas por pontes
de hidrogénio que envolvem ¢ ajudam 2 estabilizar a maioria
das biomoléculas em solugdo aquosa; (3) a distorgao dos subs-
tratos que precisa ocorrer em muitas reacbes.e (4) a necessidade
de se obter um alinhamento adequado dos grupos cataliticos
funcionais na enzima. A energia de ligaco pode ser usada para
vencer todas essas barreiras
‘A vantagem mais evidente da ligacdo dos substratos & enzi-
ma € ums grande redugio nos movimentos relativos. ou redu-
gio da entropia, desses substratos. A energia de igacio mantém
(0s sulstratos orientados adequadamente para reagir. Isso repre-
senta uma importante contribuicao & catélise, uma ver que 9s
colisdes produtivas entre as moléculas em solugdo podem ser
‘muito raras. Esse alinhamento preciso do substrato sobre « su-
perficie da enzima é devido a grande quantidade de interagdes,
fracas entre cada molécula de substrato e grupos estrategicamente
localizados na enzima que fixam as moléculas de substrato na
posigio adequada. Estudos tém demonstrado que @ restrigao do
‘movimento de dois reagentes pode produzir aumentos de velo-
‘cidade da ordem de 10°M (Fig. 8-7).
5 da velocidad
i z
ch
“ 1
CH
cele
Lo. J
Eon on
i i
a Lon ays “
j
Figura 8-7 — Aumento da velocidade por meio da reducio da en-
‘wopla, Sto representadas reaches de um éster com um grupe carbon
<2 para formar um anidhido. © grupo R @0 mesmo em cade caso. (a) Para
‘6s reaqdo bvolecula, @ corstante de veoudade € de segunda orem
‘com unidade M's. b) Quando os dois grupos reagentesestao na mes-
‘ma molécla, a reacio ¢ muito mass rapa. Para esta reacdo unimclecu-
lar, a undade de k'¢s', Dwidindo-se a cnstante de veocidade de b)
pela corstante de velocdade de (a), ter-se um aumento da velocdade
Ge cerca de 10°M (a unidade molaridade 6 resuitante 6o fato que ft
comparade uma reagio unimolecuiar com outta brmolecular). Calacan-
Corse de out modo, seo reagente em (b)estivesse presente em ua
‘oncentiagio 1M, 05 grupos reagentes se comportatam como se est
‘essem presentes em uma concentragdo 10M. Emborao reagente em
{b) apresente ie rotacdo em tres lgacOes (mostradas com setas curva)
iso ainda representa Uma substancalreducdo da ertropia em real
(@) Se 25 lgactes que gram em (h)fcam impedidas como em ie) 3
‘entopia & reduaida mais aida © a reagio exbe um aumento de veloc
ode de 10°M er relgdo 9 (a.
‘A formacao de ligacoes fracas entre o substrato e a enzima
também resulta na dessolvatagao do substrato. As interagdes en-
zima-substrato substituem a maior parte (ou até mesmo todas)
das pontes de hidrogénio existentes entre o substrato e @ dgua.
‘Acnergia de ligagio envolvendo as interagoes fracas formadas
apenas no estado de transigao da reagaoajuda a compensar termo-
dinamicamente qualquer distorcao (fundamentakmente uma re-
distribuigao de ektrons) que o substrato deva soficr para reagin,
Finalmente, a propria enzima pode sofrer mudangas con-
formacionais induzidas pelas multiplas interagdes fracas com
substrato, quando da ligacao com o substrato, Esse fato € deno-
minado de ajuste indurido, um mecanismo postulado por Da-
nie] Koshland, em 1958. © ajuste induzido serve para colocar
grupos funcionais especificos da enzima em ume posigac apro-
priads para catalisar a teagio, A mudanga conformacional tam-
bem permite a formagia ce interagdes adicionais por ligagdes
fiacasno estado de transicao, Em quaisquer desses casos. nova
conformasao apresenta propriedades cataliticas aumentadas,
Como vimos, 0 ajuste induzido é uma caracteristica comum da
ligagdo zeversivel dos ligantes as proteinas (Capitulo 7).O ajuste
induzido também € importante na interayao de quase todas as
enzimas com 0s seus substrates.
Fu
wba be spin ete
#
tle ion
open
RY HOH
L
R
Now
197
Grupos cataliticos especificos
contribuem para a catalise
Uma vex que o substrato sc liga a enzima, os grupos cataliticos
funcionais, adequadamente posicionados, auxiliam a quebra ea
formagao de ligapdes por meio de uma variedade de mecanis-
mos, incluindo a catélise Acido-base geral, a catlise covalente ©
a catélise por fons metélicos. Esses sia mecanismos distintos.
daqueles bascados na energia de ligagdo, uma vez.que geralmente
envolvem uma interacao covalente transiente com o substrate ou
‘uma transferéncia de grupos do substrato ou para 6 substrato,
Catélise dcido-base gerall, Muitas reagées bioquimicas envol-
vem a formagao de intermediarios carregados instaveis que ten-
dem a se transformar rapidamente em suas espécies reagentes.
constituintes,impedindo assim areagao (Fig. 8-8).Esses interme-
digrios carregados freqiientemente podem ser estabilizados pela
transferéncia (retirada ou adigio) de protons do substrato ou
dde um intermediario, para formar espécies cue se quebram em
produtos mais rapidamente que os reagentes. Para as reacies
ndo-cnziméticas,a transferéncia de protons pode envolver ape
nas os constituintes da dgua ou também outros doadores ou
aceptores fracos de prétons.A catélise que envolveapenas 0s ions
Produtos
Fgura 80 - Desenvolvimento desfavorival de cargas durante a quebra de uma li
'2 mesma reagdo catalizada pela quimotipsina © outras proteases, O derenvolvenento da carga ¢ des'avo-
mostrada @ hdrblise de uma ligagéo ami
10 amida que pode ser evitado pela catilise. E
idvel = pode er evtado pela doaéo de um proton peo H:0" (catélie cde especica) ou HA icatdise dcida gerll, em que HA representa um acido.
fualquee. Simiermente, a carga pode ser nevtralizada gela capture de um pioton pelo CH (catdise basica espectica) ou B: (catdlie bésica geval, em
que B: representa uma base qualquer
198
1 (H,O') ou OFF, presentes na dgua, é denominada catélise
cido-base especifica. Se a transferéncia de protons entre o in-
termedisrio ea Sgua mais rpida que a transformagio do in:
termedirio nos reagentes, o intermediario sera efetivamente
estabilirado todas as vezes que se formar. Desse modo, nenhu-
ma catilise adicional, mediada por outros receptores ou dosdo-
res de protons, ocorrerd. Entretanto, em muitos casos, apenas a
participacao da dgua nao ¢ suficiente. O termo catilise dcido-
base geral refere-se a transferéncias de protons mediacas por
coutras classes de moléculas. Para reaches nao-enzimdticas em
solugies aquosas, isso ocorre apenas quando o intermediario
instavel da reagdo se transforma nos reagentes com uma veloci~
dade maior com que os protons sio transferidos da gua ou para
2 dgua. Nessas situagdes, varios dcidos organicos fracos podem
suuplementar a égua como doadores de prétons, ov, zinda, bases
‘orgiinicas fracas podem servir como aceptoras de prétons.
Nocentro ativo de uma enzima, um certo ntimero de cadeias
laterais de aminoscides pode atuar tanto como doador quanto
‘como aceptor de protons (Pig. 6-9). Esses grupos podem ser po:
sicionados precisamente no centro ativo de uma enzima, perm
tindo a transferéncia de protons ¢ garantindo aumentos na ve-
locidade da reagao da ordem de 10° a 10°. Esse tipo de catalise
‘cote com a maioria das enzimas. De fato, as transferéncias de
protons sio as reacces bioguimicas mais comuns.
R-coon
H
RONIE
n
ROSH
Cer
ROH RHO
Oe +O
Figura §8 - Os aminodcidos em uma catilise dcido-base geral
‘Multa reacOes organicas s30 promavdas por doadores de protons (ac
os geras) ou aceptores de protons (bases geras). Os sites catalticos de
algumas enzmas contém grupos funciona de aminoscdos. como os
mestrados aqui, que podem participar no processo cataliico como doa
ores ou aceptores de prétons.
Catélise covalente. Neste tipo de catélise ¢ formada uma h
g2¢8o covalente transivéria entre a enzims ¢ 0 substrato. Const
dere a hidrdlise da tigacao quimica entre 0s grupos A eB:
ABUL A YB
Na presenca de um catalisador covalente (uma enzima com um
grupo nucleoflice X:), a reagao ocorre da seguinte forma:
ASB+Xi—p AK BOY AEA EB
Isso altera © caminho da reagdo.e resulta em catiliseapenas quan-
do 0 novo caminho tem uma energia de ativacdo menor que
aquele da reacio nio-catalisada. Assim, os dois novos pasos
precisam ser mais répidos que os da reagao ndo-catalisada. Al
sgumas cadeias laterais de aminadcidos, incluindo as da Figu-
F489, € 0s grupos funcionais de alguns co-fatores de enzimas,
podem servir como nucleéfilos na formagio de ligagbes cova
lentes com os substratos, Esses complexos covalentes sempre
sofrem rea¢Oes posteriores para regenerar a enzima livre. A liga-
cdo covalente formada entre a enzima e o substrato pode ativar
esse substrato para uma reagio subseqiiente de uma maneira
particularmente especifica para o grapo ou coenzima envolvidos.
ms
Catalise por ions metalicos. Os metais firmemente ligados
A moléculs da enzima ou captados da solucio junto com 0 subs
trato podem participar de virias maneiras na catalise. As intera-
(Ges idnicas entre um metal ligado & enzima e o substrato po-
dem ajudar a orientar o substrato para a reagao ou estabilizar os
estados de transigao carregados eletricamente. O uso de intera-
«es por ligagées fracas entre © metal e 0 substrato ¢ similar 4
utilizarao da energia de ligagao enzima-substrato ja descrita an-
teriormente.Os metais podem também mediar reagBes de ost
redugdo por meio de mudangas reversiveis no estado de
«a0 do fon metalico, Cerca de um tergo das enzimas conh
equer um ou mais ions metilicos para sua atividade catalitica
‘A maioria das enzimas emprega ume combinagao de varias,
estratégias cataliticas para aumentar a velocidade da reagao. Um
bom exemplo do uso des catilises covalente ¢ écido-base geral é
4 reacio catalisada pela quimotripsina. O primeiro ¢ a quebra
dde uma ligag3o peptidica que ¢ acompanhads pela formagio de
‘uma ligagdo covalente entre um residuo de serina da enzima ¢
parte do substrato. A reacio € acelerada pela catilise bisica geral
envolvendo outros grupos na enzima (Fig. 8-10). A reagio da
‘quimotripsina seré descrita com maiores detalhes mais adiante
neste capitulo.
mr.
EF
Figura 8-10 ~ Catalises covalente e acido-base geral. O primero pas-
$0 0 reacdo cataisada pela quenotrpsina ¢ chamado de passo da acla-
‘G20. O anupo hiro da Ser"” ¢ 0 nuceafiica em uma reacso aceera-
da pala catsise basca goral (a base & 2 cade lateral da Hes"). sso oro-
[Porcona um novo camsinho para ahidrolse da igacSo peptidca. A catahse
Somente ocorre se cada passe do novo caminho for mais rSpido que o da
reagho ndo-catalaada. A reacBo da quimotrpane esta dexrita com mas.
detahes na Fguie 8-19,
A Cinética Enzimatica como uma
Abordagem para a Compreensao do
Mecanismo de Acdo das Enzimas
Miltiplas abordagens sio comumente empregadas para estudar
‘omecanismo de agio de uma enzima purificada, O conhecimen-
to da estrutura tridimensional da protefna fornece informaches
importantese o valor dessa informagao estrutural € aumentado
significativamente pelos conhecimentos obtidos por meio da
quimica de proteina cléssica e pelos métodos modernos de mu-
tagénese sitio dirigida (mudanga da sequencia de aminodcidos
de uma proteina por meio da engenharia genética: veia Capitu-
4 29), Essas tecrologias permitem ao enrimologista exeminar
© papel individual dos aminodcidos tanto na estrutura da enzi
ma como na sua aco catalitica, Entretanto, a abordagem cen-
tral para estudaro mecanismo de umareagio catalisada por uma
cenzima é determinat 2 velocidade da reagao ¢ como ela se altera
‘emfungdo de mudancas nos parimetros experimentais, uma dis-
ciplina conhecida como cinética enzimatica. Essa é 2 aborda
‘gem mais antiga para 2 compreensio des mecanismos enzima-
tices ¢ continua sendo muita importante até hoje. 0 que segue é
uma introdusdo bésica& cinetica das reagses catalisadas por en-
2imas. Para um estudo mais avancado devem ser consultadas as
referéncias citadas no final do capitulo,
A concentracao do substrato afeta a velocidade
das reages catalisadas por enzimas
‘Um dos principais fatores que afetam a velocidade de uma rea
80 in vitro, catalisada por ume enaima purificada, é a concen
tragao do substrato, [S]. Entretanto, estudar os efeitos da con-
centeagio do subsirato & complicado pelo fato de [S] variar du-
rante © curso de uma deda reagao, & medida que o substrato €
convertido em produto, Uma abordager simplificada em expe-
simentos cinéticos é medir a velocidade inicial da reagio, desig.
nada por Vq, quando [S] € geralmente muito maior que a concen:
tracdo da enzima, [F|. Assim, se 0 tempo de reacio for suficien-
temente curto, as mudang2s na {S] serio despreziveis, portanto
2 |S] pode ser considerada constante.
© efeito em Ve provocado pela variagio da [S}, quando a
concentracio de enzima € mantida constante, esté mostrado na
Figura 8-11, Em concentracées relativamente baixas de substra-
to, Vi aumenta quase linearmente com o aumento da [S]. Em
altas concentragies de substrato, Vp aumenta cada vez menos
em resposta aos aumentos da (S|. Finalmente, é alcanzado um
ponto acima do qual ocorrem aumentos insignificantes de Vi, &
medida que a [S] aumenta, Esse patamar atingido para tais va-
lores de Vo é muito proximo de velocidade maxima, Vmax
(© complexo ES é fundamental para a compreensdo desse
comportamento cinético, ji que ele foi o ponto de partida para a
Aiscussio da cavdlise enzimatica, © perfil cinético apresentado
na Figura 8-11 levou Victor Henri, seguindo as diretrizes de
Wirtz, « propor, em 1903, que « combinagao de uma enzima
com a mokcula de seu substrato para formar 0 complexo ES €
uum passo obrigatério na catdlise enzimética. Fm 1913 essa idéia
foiexpandida em uma teoria geral da ago das enzimas por Leo”
nor Michaelis e Maud Menten. Eles postularam que inicialmen
te a enzima se combina reversivelmente com o substrato para
formar 0 complero enzima-substrato, em um passo reversive!
relativamente répido:
Esse
BS 6)
Em uma segunda etapa lenta, o complexo ES entio se quebra
liberando a enzima livre, o produto da reagio, P:
pcteerr ws
‘Como a segunda reag3o & mais lenta (Eq. 8-8), ela limita a velo-
lade da reagao enzimatica. Assim, a velocidade da reagao erzi-
mmitica deve ser proporcional 3 concentragdo da substincia que
reage no segundo passo, isto & ES.
Em qualquer instante de uma reacao catalisada enzimatica
‘mente, enzima existe em duas formas, ni ligada ao substra-
0,04 forma livre E, a forma ligada 40 substrato, ES. Em baixas
{S],amaior parteda enzima esté na forma livre E. Nessascondi-
Ges, a velocidade da reagdo ser proporcional & [S] porque ©
equilibrio da Equacao 8-7 € deslocado na direcao da formato
y(n)
g
4
2
2
‘Concentracao de substrato, [S] (mM)
Figura 6-11 ~ Efeito da concentragéo de substrato na velocidade
inielal de uma reacéo eatalisada enzimaticamente, Por me deste
tipo de grafico, 2 Vnys pode ser determinada apenas aprosirradamente,
|sh0 porque Ys 3 apraxima mas nunca chega a abngir Vnw. A concentra
{0 de substrazo, em que a Vy ¢ a retade da Vras,# Ky, a constante de
Michaelis-Menten. & concentiacao da enzima em Um experimento coma
‘esse ¢ getalmente t3o baixa que [3] >> [E] mesmo quando [S| ¢ descta
como pequens ou relativamente pequena, As uridades sao tipicas para
reactes cataisadas engimaticarmerte e apresertadas apenas para iustrar
lo signdicade Use [S] (note que 2 curva descreve parte de uma hiptrbole
‘retanguiay, com uma das assintotas em Vaq: Se 3 curva continuasse
eb se aproximaria de ume assiniota
Leonor Michzelis
(1875-1949)
‘Maud Menten
(1879-1960)
dde mais ES, 8 medida que a |S| sumenta. A velocidade inicial
maxima da reaglo (Vue) sera atingida quando praticamente
todas as moléculas da enzima estiverem na forma do complexo
ES ca concentragso da enzima livre E for significativamente pe-
‘quena, Nesss condigdes, a enzima esta“saturada’ com seu subs-
trato ea velocidade da reacio no aumenta mais com novos au-
mentos da (S]. Essa condigdo existiré sempre que a [] for sufi-
ccientemente alta para manter todas as moléculas de enzima na
forma combinada com o substrato, ES. Em seguida,o complexo
ES transforma-se no produto P e a enzimaé liberada para cata:
lisar a transformagio de outra moléeula de substrato. O feito
dle saturagao € uma caracteristica que distingue os catalisadores
enzimaticos e & 0 responsivel pelo patamar observado na Figura
8-11, Essa condigio de saturagii existird sempre que [S] for su-
ficientemente alta,
Quando a enzima & misturada com um grande excesso de
substrato, existe um periodo inicial, o estado pré-estacionério,
‘no qual a concentracdo de ES aumenta. Esse periodo ¢ muito
curto para poder ser facilmente observado. A reacio atinge rapi
damente 0 estado estaciondrio, em que a [ES| (ea concentragao
199
200
de quaisquer outros intermediarios) permanecerd praticamente
constante durante 0 decorrer de certo tempo.O conceito de es-
tado estaciondrig foi introduzido por G.E. Briggs ¢ J.B.S, Halda-
neem 1925. A velocidade medida Vp geralmente refleteo estado
estacionério (mesmo considerando-se que Voestejalimitada aos
tempos iniciais da reacdo) ¢ a anilise dessas velocidades iniciais
& chamada de cinética de estado estacionério,
A relacdo entre a concentracao do substrato
@ a velocidade da reagSo pode ser expressa
quantitativamente
A curva que expressa a relacio entre [S] e Va(Fig. 8-11) tem a
mesma forma geral paraa maioria das enzimas (ela se aproxima
de uma hipérbole retangular) que pode ser expressa algebrica-
mente pela equagio de Michaelis-Menten. Michaelis ¢ Menten
erivaram essa equacdo partindo de sua hipdtese bésica de que
© passo limitante da velocidade nas reacSes enzimaticas seria «
quebra do complexo ES para formar 0 produto e a enzima livre.
Acquacdo é
(89)
Os termos importantes s4o [S]. Vo. Vaux € uma constante cha-
mada de constante de Michaelis, Ky. Todos esses termos sdo fia-
cilmente medidos experimentalmente.
Nés desenvolveremos aqui a logica bisica e os passos algé-
bricos da dedugio moderna da equasao de Michaelis-Menten,
que inclui a consideragio do estado estaciondrio introduzide
por Briggs © Haldane, A dedusdo comeya com as duas etapas
‘bdsicas envolvidas na formagio e na quebra de ES (Eqs. 8-7 €8-
8). Nos primeiros momentos da reagio, a concentracio do pro-
duto, [P], € negligencitvel , para simplificar, assumiremos que
‘k,(que descreve a reacio reversa de P para S) pode ser ignora-
da. Essa consideragiio nao é critica mas simplifica nossa tarcfa,
Assim, a reagao geral se reduz a
pescteesctersr (00)
Vo determinada pela quebra de ES para formar o produto, que
or sua vez é determinado pela [ES]:
Vo= IslES|
Como « [ES] na Equasie 8-11 nio € facilmente medida experi-
‘mentalmente,¢ preciso encontrar uma expressdo alternativa para
[ES]. Primeiro, introduziremos © termo [E] que representa a
‘concentracdo total da enzima (a soma das concentrac0es da en-
ima livre e da ligada ao substrato). Desse modo, a concentragio
> [5] € 0
temme [5] n@ denominacor da equagdo de Mchaelis-Mertten (Eq. 8-9)
tomas insignficante. A enuacso pode ser simplificade para Vo = Vous
ISbk,. @ Vy pasta a tor uma dependéncia linear ern fungao da [S], coma
pode ser cservado nesta figura. Er [S]elevadas, onde |S] >> Ky, 0 tor
Imo Kr. na denernineder da equegdo de Michaelis Merten torna:se insig-
hifcante e a equacao pode ser simplfcada pata Yo = Vrax 80 & consis
teste com o patamar cbservado nas [5] elevades.Portento, a equaca0 ce
Michaels-Menten & consstente com a depend2ncia observada de Vem
ralacio a [S] A forma da curva é definida pelos termes Yru/Kn, quando
a[5}@ pequena, ¢ apenas for Vue quanda a {5} alta
201
‘A equagdo de Michaelis-Menten (Eq. 8-9) pode ser trans-
formada algebricamente em formas que sao mais uteis para a
determinagio pritica de Kye Vaw, (Adendo 8-1) e, como desere-
vveremos posteriormente, na andlise da agdo de inibidores (veja
Adendo 8-2).
Os parametros cinéticos sdo usados
para comparar as atividades das enzimas
importante distinguir entre a equacae de Michaelis-Menten &
‘omecanismo cinético especificn em que ele se baseia. A equagdo
descreve 0 comportamento cinético da grande maioria des enzi-
‘mas, todas as enzimas que exibem uma dependéncia hiperbo-
lica de Vo em relagioa |S] sio denominadas enrimas que segue
a cinética de Michaelis-Menten, A regre pratica de que Kn= (S]
quando Vo=} Vy, (Eq, 8-23) € vilida para todas.as enzimas que
seguem a cinética de Michactis-Menten (a maioria das excegdes
a cinética de Michaelis-Menten sito as enzimas reguladoras, dis-
ceutidas no final deste capitulo). Entretanto, a equago de Mi-
chaelis-Menten nao depende do mecanismo de reagio relat
mente simples, de duas etapas, proposto por Michaelis e Menten
(Eq, 8-10), Muitas enzimas que seguem a equagao de Michaelis:
‘Menten tém mecanismos de reacdo muito diferentes entte si, ¢
enzimas que catalisam reagdes com seis ou oito etapas identifi
clveis freqdentemente exibem 0 mesmo comportamento ciné
tico de estado estaciondrio. Mesmo considerando que a Equa:
Adendo 8-1 | Transformacées da equacao de Michaelis-Menten:
© grafico dos duplos reciprocos
A equaciio de Michaelis-Menten
Val
1 n+ (SI
pode ser transformada algebricamente em equacdes,
zitir uma determinagao mais acurada de Vas ¢uUe
pode ser obtida apenas aproximadamente pelo gré-
fico Vo versus [S] (veja Fig. 8-12).
Outras transformagoes da equagio de Michae~
lis-Menten tém sido deducidas, cada ume apresen-
ue sio mais iteis no tratamento grifico dos dados
expetimentais, Uma transformagao comum ¢ obti-
da simplesmente invertende-se 0s dois lados a
equacio de Michaelis-Menten:
A _ Ko t181
Vo nal]
Separando-se 03 componentes do numeradot do
lado direito da equagio obtém-se
ieee SE
Vy Val] Vas)
que pode ser simplificeda para:
Ava Say,
Va Vinal]
Vous
Esta forma da equacio de Michaelis-Menten & cha-
made de equegdo de Linewerver-Burk. Para as en-
zimas que obedecem a equagio de Michaelis-Men-
ten, 0 grafico de I!Va versus 1/(S] (gréfico des “da
plos reesprocos” de Vs versus (S] que usemos até
agora) é representado por uma linha reta (Fig. 1).
Essa linha tem uma inclinego de Kiy/Vins tm n=
tercepto de 1/V ny n0 eivo de 1/Vp € um intercepto
de—I/Ky no eixo de 1/[S]. A representagaa dos du-
plos reciprocos, também chamada representagao de
tando alguma vantagem particular na anilise de
dados de cinética enzimitica (veja problema 1,
ig, 229).
‘0 grifico dos duplos reciprocos para velocida-
des de reagdes enzimiticas ¢ muito itil na diferen-
ciagae de diferentes mevanismos de reagao enzim’-
tica (veja Fig. 8-14) e na andlise de inibidotes de
cenzimas (veja Adendo 8-2).
Figura 10 gr8fco dos duploerecipracos de linewea-
ineweaver Burk, tem a grande vantage de per ver Burk
202
(ao 8-25 € verladeira para muitas enzimas, a magnitude e o sig-
nificado real de Vin Ky podem variar de uma enzima para
outra. 1550 € una limitagao importante do modelo do estado
estaciondrio para a cinética enzimitica, Vay. ¢ K, sio parime-
tros que podem ser obtidos experimentalmente para qualquer
entima, mas mesmo assim eles fornecem pouca informagao so-
bre o ntimero, a velocidade ou a natureza quimica das etapas
discretas da reagdo, Nao obstante, a cinética de estado esta-
cionério representa a linguagem-pedrao pela qual as eficigncias
> ke entdo
Ky = ki/ky. Em outros c4s0s, k, ¢ k.; s8o compardveis ¢ Kmy per
manece como uma fungae mais complexa das trés constantes
de velocidade (Eq. 8-24). Nesse caso, a equagao de Michaelis.
Menten € 0 comportamento carectertstico da seturacdo da en-
vima ainda sio valides, mas 0 Ky nao pode ser considerado
come uma medida da afinidade da enzima pelo substrato..
da mais comtns sio 05 casos em que a reagio ocorre por meio
de etapas miltiplas apés a formacio do complexo ES. O Ky
enlao se torna uma fungao muite complexa de muitas constan-
tes de velocidade
‘Tabela £-6 ~ Valores de Kq para algumas enzimas e substratos
Enzima Substrato CACO)
Catalase HO; 35
Hexoquinase (cerebro) ATP. oa
Deglicose 0.05
Defrutose 15
Aricrase carbénica HeO5 26
Quimotripsna Gicitvosnigicina 108
‘Nbenzoitirosinamida 25
Pegalactosidase DHactose 40
“Tiecnina desicratase Lareorina 50
A Vy também varia muito de ums enzima para outra. Se
uma enzima reage de acordo com o mecanismo de duas etapas
Michaelis-Menten, Vx & equivalente a ka(F,), onde ks & a etapa
limitante da velocidade, Entretanto, 6 niéimero de etapas de rea~
so € a identidade da(s) etapa(s) limitente(s) da velocidade
podem variar de enzima para enzima. Por exemplo, considere
uma situagio muito comum na qual a liberagio do produto,
EP > E +R limitante da velocidade. Nos instantes iniciais da
reagio (quando a [P] ébaixal,« rea;ao global pode ser descrita
pelo esquerna
ke
Ee Stes ee ep es (23)
‘Neste caso, a maior parte ds enzima estd na forma EPem con
‘goes Ge saturagdo © Ving = Ks[Eq. E wil definir uma constance
mais geral, kuw» pata descrever a velocidade limitante de qual-
‘quer reagio cataisada por uma enzima em condigoes de satura
‘G20. Se existirem varias etapas na reacao enzimatica e uma delas
for visivelmente limitanteda velocidad, k.« éequivalente dcons-
tante de velocidade da etapa limitante, Para a reagao éa Equagio
8-10, kan = ke, Pera a reagao da Equacao 8-25, kx = ky. Quando
varias etapas sio parcialmente limitantesda velocidade, kx pode
se ransformar em uma fungae complexa de varias das constantes
de velocidade que definem cada etapa individual da reagio, Na
Vaau![Ex] € a Equagio 8-9
se transforma em
(8.28)
A constante k.,, & uma constante de velocidade de primeirs or-
dem cuja unidade € 0 reciproco do tempo. Bla também é chama-
da de numero de renovacao e é equivalente ao ntimero de molé-
ceulas do substrato convertidas em produto por uma tinica mo
lecula da enzima, em uma dada unidade de tempo, quando a
‘enzima esta saturada pelo substrato, Os miimeros de renovagio
«le varias enzimas so apresentados na Tabela 8-7.
Tabela 8-7 ~ Numero de renovagio (kay) de algumas enzimas
Enzima Substrato Ken ©)
Catalase 1.0; 0,000,000
Anidrase carbonica HCO; 400.000
Aceticolinesterase Acetiatina 140.000
lactamase Benailpeniciina 2.000
Fumarare Fumarate 200
Proteina Rach (uma ATPase) ATP a4
(Os parametros cinéticos ky € Ky geralmente sio titeis para
‘o estudo e a comparagio de diferentes enzimas independente-
‘mente de seus mecanismos de reagao serem simples ou comple
x0s.Cada enzima apresenta valores dtimos de ka € Ke, que refle-
tem o ambiente celular a concentragio do substrato nermal-
mente encontraco in vivo pela enzimae a quimica da reagio que
esté sendo eatalisada
Os paramettos ks € Kr, permitem avaliara eficiencie cata-
Iitica das envimes, mas esses parametras isoladamente sto in-
ntes para essa tarefa, Duas enzimas que cat
‘Goes diferentes podem ter a mesma k.. (miimero de renovagao),
embora as velocidades das reagbes nio-catalisadas possam ser
diferentes. Dessa forma, o aumento da velocidade provocado
pela enzima pode ser muito diferente. Experimentalmente. 0
Kp de uma enzima tende a ser similar a concentragio celular do
seu substrato, Uma enzima que atua sobre um substrato pre-
sente em uma concentrarao muito baixa no interior da céhula
tenderd a ter um Ky, muito menor que aquela que atua sobre
‘um substrato que é mais abundante,
‘Armelhor maneira de se comparar eficigncia catlitica de
diferentes enzimas ou onamero de renovacdo de diferentes subs-
tratos para uma mesma enzima 6 analisar a telagio Kay/Ky para
as duas reagoes. Esse parametro, que algumas vezes é denomina-
do constante especifica, é @ constante de velocidade para acon-
yersao de E + 5 em E + P Quando [5] << Kp, a Equacao 8-26 se
redus & forma
Es)
27)
Neste caso, como V depende da concentragao de dois reagen-
tes, [F,] e [S], ela é uma equagio de velocidade de segunda or-
dem e 2 constante ka/Km € uma constante de velocidade de
reazio de segunds ordem com unidades M's“, Bxiste um li-
nite superior para hea/ Ky» imposto pela velocidade com que E
eSsedifundem em uma solucdo aquosa, Esse limite, controla-
do pela difusao, 6 da ordem de 108a 10° Mle, e muitas enai-
mas apresentam 0 valor de Kas/Ky proximo a esse intervalo
(Tabela 8-8). Tais ensimas s40 consideradas cataliticamente
perfcitas, Note que diferentes valores de Ky: € Km postem produ
tir relagdes maximas.
Muitas enzimas catalisam reagdes ./
que envolvem dois ou mais substratos
Jévimos come a [S] afeta a velocidade de uma reagao enzimiti
a simples ($ > P) na qual participa apenas uma molécula de
substrato, Entretanto, em muitas reagbes enzimaticas, duas (al-
‘gumas vezes mais deduas) moléculas de substratos diferentes se
ligam a enzima e participam de reacdo. Por exemplo, na reagao
‘atalisada pela hexoquinase, o ATP e a glicose sio as moléculas
ce substrato & 0 ADP € a glicose-5-fosféto sto os produtos:
ATP + glicase —> ADP + glicose-o-fosfato
As velocidades dessas reagoes com dois substratos também po:
em ser analisadas pela abordagem de Michaelis-Menten. A he-
xoquinase tem um K,,caracteristice para cada um dos seus dois
substratos (Taela 8-6).
‘As reagdes enzimaticas com dois substratos ususlmente en
vlvem a transferéncia de um étomo ou um grupo funcional de
um substrato para 0 outro. Tais reagbes ocorrem por meio de
tum ou vérios caminhos diferentes. Em alguns casos, embos os
sabstratos estéo figados 4 enizims ao mesmo tempo, em algum
instante da rea¢io, formando um complexo terndtio nie-cova-
lente (Fig. 8-13a). Esse compiexo pode ser formado pela ligagao
dos substratos em uma seqiléncia zo acaso ou em uma ordem
epecifica. Nenhum complexo ternério ¢ formado se 0 primeiro
substrato é convertido em produto e se dissocia antes ca ligagdo
do segundo substeato, Um exemplo ¢ 0 mecanismo chamado
pingue-pongue ou de dupla-tioca (Fig. 8-13b). A cinética do
estado estaciondrio freqiientemente pode ajudar na distingio
ntre essas porsibilidades (Fig. 8-14)
Figura 8-13 - Mecanismos comuns para raaghes com dois subs-
tratot catalisados enzimaticamente. Em (a) 3 enzrna © ambes 05
substrates se juntar para fermar um complexo ternario, Em uma 2sso-
Cdlagao ordenada, 0 substrata 1 deve ser ligaco antes, para que o subs
tate 2 possa se ligar produtvamente. Em uma associaga0 20 acaso, 0s
substrétos podem se ligar em qualgver order, Em (b), um comglexo
lenzime-substrato se forma, um produro ¢ formado, a encima modifica-
dda forma um segundo comelesa com uma molécula de un auto subs-
trate, ¢ um segunca produto & fermado, regenerando a encima ive, O
substrate 1 pode transfenr um grupo funcional para @ endima (ork
nando a forma modiicada covalentemente, £), que © subsequent-
mente transferiéa para 0 substrato 2. fsse @ 0 ‘mecanismo chamado
pinque-pongue ou de duple-toca
‘A cintica de estado pré-estaciondrio pode fornecer
evidencias para etapas especificas da reacdo
‘A cinética enzimitica foi introduzida como um método impor-
tante para estudar as etapas deuma reacdo enzimatica,bemcomo
foram mencionadas as imitagdes dos pardmetros cinéticos mais
comuns que fornecem tal informaga0. Os dois parametros ex-
perimentais mais importantes fornecidos pela cinética de esta-
do estaciondrio so © key € © Ku/Kr. As variagdes desses pari
‘metros com a variagao do pH e da temperature podem fornecer
informagées adicionais a respeito das etapas de uma teagio. No
caso de teagdes com dois substratas,a cinética do estado estacio-
nario pode ajudar a determinar se durante 2 reagdo € formado
um complexo ternirio (Pig. 8-14), Um quadro mais completo
geralmente requer métodos cineticos mais sofisticados, que vo
além dos objetivos de uum texto introcutério, Assim, serd intro
duzida, de forma muito breve, uma des mais importantes abor
degens para se estudar o mecanismo de uma reacio, a cinética
de estado pré-estacionério.
‘abela £8 — Enzimas nas quals 0 valor de hua/Kn esté préximo do limite controlado pela difusdo (de 10% a 10° Mrs")
tncina sarees i 5 a
Acetilcolinesterase Acetileolina 414x108 ox 10% 1.6 x 108
Anidrase carbinica Oz 1x10? 12% 107 83x10"
ros ie ae 8
Catalase Hj, 42107 WwW 4x10"
— Seth soicle 2x10 apne
Malato 9x10? 25x 107 3.6 x 10"
sci seria aise zai sit
Fonte Fest, A (1898). Structure and Mechanism Pretan Seance, p. 166, Freeman and Company, Newvork
204
isi =
Figura #-14 — Andlise cinética do estado estacionario para rea-
es com dois substratos, Nestes graficos dos cuplos tedprocos (vale
‘Adendo 8-1). a concentracdo do substrato 1 & vardvel, enquanto 2 con
centrazaa do substrata 2 6 mantida constanto, x0 6 repetido pata 3
nos valores de [5], gerando viras etas seperadas, A intersegho desias
retas indige a formacéo de um complesa ternéria na reacie (a); retas
paralels indicam que 2 reacao ocorre por uma via do tipo pingue-pon-
{gue ou dupla-traca fb)
Uma descrisao completa de uma reeg2o catalisada por enzl-
‘mas requer medidas diretas das velocidades das etapas indivi-
duais da reago, como por cxemplo a medida da associagao da
enzima e com o substrato para formar o complexo ES, & duran-
te ocstado pré-estaciondrio que as velocidades de muitas etapas
dda reagdo podem ser medidas independeniemente. As condigoes
da reacio sao ajustadas para facilitar a medida dos eventos que
ccorrem durante a transformagée de uma tinica molécula de
substrato, Uma ver cue a fase de estaco pré-estacionaio em ge-
ralé muito curts, freqilentemente sio necessarias técnicas espe-
calizadas para efetuar uma mistura muito répida dos reagentes
e aamostragem que se segue. Um dos objetives é a obtencao de
‘um quedro completo e quantitative des mudangas de energia
durante a reagio. Conforme mencionado anteriormente,a velo-
cidade das reagGes e 0 equiltbrio estio relacionados com as mu-
dangas de energia livre que ocorrem durante a reagio, A medida
da velocidade das etaps individuais de uma reagdo revela como
‘aenergia é usada por uma enzima especifica, © que representa
‘um componente importante do mecanismo global da rea¢éo.
Em um grande ntimero de casos jt é possivel medir a velocidade
cc cada uma das etapes individuais de ume teagao enzimstica
‘que apresenta varias etapas, Alguns exemplos da aplicagao da
cinética de estado pré-estacionsrio estdo incluides nas deseri-
goes de enzimas especificas, ainda neste capitulo.
As enzimas s60 sujeltas & inibigso
s inibidores das enzimas sdo agentes moleculares que interfe-
rem com a catdlise, diminaindo ow interrompendo as reagdes
enzimaticas. As enzimas catalisam virtualmente todos os pro-
cessos celulares e nao é surpreendente que os inibidores das en-
sci ekg ates bu tala ane eas anemia
conhecidos. Por exemplo, a aspirina (acetisaliclato) inibe aen-
ima que cataliss 0 primeiro passo na sintese das prostaglan
nas, compostos envolvidos em muitos processos,incuindo al
guns que produzem a sensacio de dor. © estudo dos inibidores
das enimas tem fornecido valivas informagies a respelto dos
rmecanismos enzimticos e tem ajudado a definir algumas vias
smetabdlicas, Existem duas grandes classes de inibicores enzi-
infticos:reversives ireversiveis,
A inibicdo reversivel pode ser
competitiva, incompetitiva ou mista
Um tipo comum de inibidor reversivel é chamado competitivo
(Fig. 8-15a). Um inibidor competitive compete com o substra-
to pelo sitio ativo da enzima, Enquanto o inibidor (1) ocuparo
i @
r@
(a) Inicio competivive
res > esse
k@2e
la
&
(b) Inibicdo incompettiva
BS BBP
1
H+s == ESL om >
(© Inibiqao mista
Figura 8-15 ~ Tr8s tipos de inlbicdo revarsivel. a} Os initidores P entio um anticorpo que
se liga fortemente ao andlogo do estado de transi-
«0 pode catalisar S~» POs anticorpos {imunoglo-
bbulinas: veia Fig. 7-23) sio os principais participan-
tes da resposta imunolégica, Quando um andlogo
do estado de transigio € empregado como uma pro-
teine ligante de epitopo para estimular a produgio
de anticorpos. os anticorpos que se ligam a ele si0
Você também pode gostar
- Aula 2 - Atomos Moleculas e Ions (Modo de Compatibilidade) Quimica GeralDocumento62 páginasAula 2 - Atomos Moleculas e Ions (Modo de Compatibilidade) Quimica GeralmayaramagdaAinda não há avaliações
- Aula 6 - Estequiometria Moodle Quimica GeralDocumento42 páginasAula 6 - Estequiometria Moodle Quimica GeralmayaramagdaAinda não há avaliações
- Aula 4 - Propriedades Periódicas Dos Elementos (Modo de Compatibilidade) Quimica GeralDocumento46 páginasAula 4 - Propriedades Periódicas Dos Elementos (Modo de Compatibilidade) Quimica GeralmayaramagdaAinda não há avaliações
- Aula 5 - Reações em Solução Aquosa (Modo de Compatibilidade) Quimica GeralDocumento39 páginasAula 5 - Reações em Solução Aquosa (Modo de Compatibilidade) Quimica GeralmayaramagdaAinda não há avaliações
- Aula 1 - Conceitos Fundamentais de QuímicaDocumento41 páginasAula 1 - Conceitos Fundamentais de QuímicaFabio de XangôAinda não há avaliações
- Aula 3 - Estrutura Eletrônica Dos Átomos (Modo de Compatibilidade) Quimica GeralDocumento61 páginasAula 3 - Estrutura Eletrônica Dos Átomos (Modo de Compatibilidade) Quimica GeralmayaramagdaAinda não há avaliações
- Aditivos PolimerosDocumento36 páginasAditivos PolimerosmayaramagdaAinda não há avaliações
- Aula1 Fisica1Documento8 páginasAula1 Fisica1mayaramagdaAinda não há avaliações
- Nomenclatura de Hidrocarbonetos RamificadosDocumento28 páginasNomenclatura de Hidrocarbonetos Ramificadosdougstardoug22085999Ainda não há avaliações
- Estabilidade Dos Sistemas ColoidaisDocumento25 páginasEstabilidade Dos Sistemas ColoidaismayaramagdaAinda não há avaliações
- Aula1 Fisica1 PDFDocumento8 páginasAula1 Fisica1 PDFmayaramagdaAinda não há avaliações
- Capítulo 2 Água Tampão LehningerDocumento19 páginasCapítulo 2 Água Tampão LehningermayaramagdaAinda não há avaliações
- Análise Da Água SlidesDocumento42 páginasAnálise Da Água SlidesmayaramagdaAinda não há avaliações
- ColoidesDocumento5 páginasColoidesthiagomaddAinda não há avaliações
- Aula 3 e 4 MineralogiaDocumento38 páginasAula 3 e 4 MineralogiamayaramagdaAinda não há avaliações
- Aula 1 Calendarios Sanitarios para Bovinos de Corte PDFDocumento36 páginasAula 1 Calendarios Sanitarios para Bovinos de Corte PDFmayaramagdaAinda não há avaliações
- Introdução À CromatografiaDocumento71 páginasIntrodução À CromatografiamayaramagdaAinda não há avaliações
- Aula 5 Rochas MetamórficasDocumento28 páginasAula 5 Rochas MetamórficasmayaramagdaAinda não há avaliações
- Relatorio ONUDocumento8 páginasRelatorio ONUEduardo CazonAinda não há avaliações
- 02 Água - Recurso Natural, Finito e Estratégico PDFDocumento21 páginas02 Água - Recurso Natural, Finito e Estratégico PDFottoramosAinda não há avaliações
- Integrais MultiplasDocumento52 páginasIntegrais MultiplasmayaramagdaAinda não há avaliações
- Capitulo Cadeia Transportadora de EletronsDocumento25 páginasCapitulo Cadeia Transportadora de Eletronsmayaramagda100% (1)
- Ign2004 4Documento24 páginasIgn2004 4Andreas GringsAinda não há avaliações
- Primeiros Socorros IFRSDocumento59 páginasPrimeiros Socorros IFRSIsabelle MayaraAinda não há avaliações
- Tecido NervosoDocumento21 páginasTecido Nervosoju mAinda não há avaliações
- A Influencia Dos Elementos Da Astrologia Do Visagismo - 20130829000604Documento21 páginasA Influencia Dos Elementos Da Astrologia Do Visagismo - 20130829000604spawn2006Ainda não há avaliações
- Aspectos Clínicos e Fisiopatológicos Da Rabdomiólise Após Esforço Físico Intenso PDFDocumento10 páginasAspectos Clínicos e Fisiopatológicos Da Rabdomiólise Após Esforço Físico Intenso PDFUrias Mendes de AquinoAinda não há avaliações
- 3 Série Geografia Aluno 4ºbiDocumento33 páginas3 Série Geografia Aluno 4ºbiVitória MartinsAinda não há avaliações
- Alcoolímetro Alcolock V3Documento3 páginasAlcoolímetro Alcolock V3TECNIQUITELAinda não há avaliações
- Treinamento - Como Elaborarum Projeto de Plantio Compensatório - 23-10-2014Documento32 páginasTreinamento - Como Elaborarum Projeto de Plantio Compensatório - 23-10-2014Hélio Cavalhéri Jr.Ainda não há avaliações
- Fungos Produtores de Micotoxinas em AlimentosDocumento53 páginasFungos Produtores de Micotoxinas em AlimentosYasodhara CapellaAinda não há avaliações
- Carta Argumentativa - Literatura FamininaDocumento2 páginasCarta Argumentativa - Literatura FamininaIsabella Unterrichter RechtenthalAinda não há avaliações
- 3 Poison ThroneDocumento360 páginas3 Poison ThronePamela MartinsAinda não há avaliações
- 2 Riscos em Instalações e Serviços Com EletricidadeDocumento34 páginas2 Riscos em Instalações e Serviços Com EletricidadeReginaldo Sant AnaAinda não há avaliações
- A Circulação Pulmonar e A Circulação SistémicaDocumento2 páginasA Circulação Pulmonar e A Circulação SistémicaVania FariaAinda não há avaliações
- Respiração Celular PDFDocumento4 páginasRespiração Celular PDFJuliana Fernanda100% (1)
- Guia de Plantas Medicinais e EnergéticasDocumento30 páginasGuia de Plantas Medicinais e EnergéticasFlávia rocha100% (1)
- Mapa AstralDocumento43 páginasMapa Astralvitoriarocha0702Ainda não há avaliações
- Resumo de Anatomia - Fernando FelixDocumento266 páginasResumo de Anatomia - Fernando FelixCarlos Vinícius JorgeAinda não há avaliações
- Questionário Parte 2 Clado Das ComelinídeasDocumento2 páginasQuestionário Parte 2 Clado Das ComelinídeasnaidooAinda não há avaliações
- Fatores Interferentes Da Absorção e Vias de AdministraçãoDocumento12 páginasFatores Interferentes Da Absorção e Vias de AdministraçãoregianeAinda não há avaliações
- Teste Reino PlantaeDocumento3 páginasTeste Reino PlantaeArlanjoAinda não há avaliações
- ANATOMIA-SECCIONAL-E-POR-IMAGEM-1 - Prof LuizDocumento32 páginasANATOMIA-SECCIONAL-E-POR-IMAGEM-1 - Prof LuizLuiz BatistaAinda não há avaliações
- CoraçãoDocumento27 páginasCoraçãoLeticiaAinda não há avaliações
- Coprocultura Protocolos Parte 2Documento5 páginasCoprocultura Protocolos Parte 2GaabfcAinda não há avaliações
- Visão Psicanalítica Do Filme CoralineDocumento5 páginasVisão Psicanalítica Do Filme CoralineLucas100% (1)
- Dinos SaurosDocumento38 páginasDinos SaurosAndreAinda não há avaliações
- Bizu QuestoesDocumento4 páginasBizu QuestoesMariAinda não há avaliações
- TP2 V de Gowin 1 - Extracção DNA ProfDocumento2 páginasTP2 V de Gowin 1 - Extracção DNA ProfRenata Rosário100% (2)
- Roficiência Motora em Crianças Normais e Com Dificuldade de Aprendizagem - Estudo Comparativo e Correlacional Com Base No Teste de Proeficiência Motora de Bruininks - OseretskyDocumento16 páginasRoficiência Motora em Crianças Normais e Com Dificuldade de Aprendizagem - Estudo Comparativo e Correlacional Com Base No Teste de Proeficiência Motora de Bruininks - OseretskyAndreia SilvaAinda não há avaliações
- O Guia Das Criaturas MágicasDocumento92 páginasO Guia Das Criaturas MágicasF. Gabriel F. CortezAinda não há avaliações
- SANARFLIX - Fisiologia Dos Órgãos Sensoriais (Visão e Olfato)Documento35 páginasSANARFLIX - Fisiologia Dos Órgãos Sensoriais (Visão e Olfato)laviAinda não há avaliações
- Questões Sobre Funções InorganicasDocumento18 páginasQuestões Sobre Funções InorganicasJessica CorreiaAinda não há avaliações