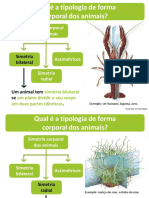
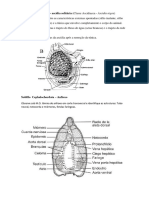
Você também pode gostar
- Protocolo de Nódulo Tireoidiano No Adulto 07 de Agosto2Documento9 páginasProtocolo de Nódulo Tireoidiano No Adulto 07 de Agosto2Valdir VidalAinda não há avaliações
- AnticoncepçãoDocumento97 páginasAnticoncepçãobaleeirodaianyAinda não há avaliações
- Rec 38 ManejoTosseDocumento10 páginasRec 38 ManejoTosseenzomontresolAinda não há avaliações
- Manejo Paciente Diarreia CartazDocumento1 páginaManejo Paciente Diarreia CartazLayane TérciaAinda não há avaliações
- Forum Franca Final v2Documento41 páginasForum Franca Final v2enzomontresolAinda não há avaliações
- Consenso RGEDocumento17 páginasConsenso RGEaureopzAinda não há avaliações
- Os Macrolidos para Alem Do Seu Efeito AntibioticoAna SofiaDocumento32 páginasOs Macrolidos para Alem Do Seu Efeito AntibioticoAna SofiaenzomontresolAinda não há avaliações
- Manual ATBDocumento41 páginasManual ATBenzomontresolAinda não há avaliações
- Afecções Da AortaDocumento31 páginasAfecções Da AortaenzomontresolAinda não há avaliações
- A Nuvem Sobre o Santuário Karl Von Eckartshausen - 41Documento41 páginasA Nuvem Sobre o Santuário Karl Von Eckartshausen - 41Fcx020781Ainda não há avaliações
- Sindrome Do Desconforto Pulmonar Do AdultoDocumento14 páginasSindrome Do Desconforto Pulmonar Do AdultoenzomontresolAinda não há avaliações
- TradDocumento22 páginasTradMarcos Carvalho LopesAinda não há avaliações
- KEVIN TRUDEAU - Curas Naturais Que Eles Não Querem Que Você Saiba - LIVRO-576 PágDocumento96 páginasKEVIN TRUDEAU - Curas Naturais Que Eles Não Querem Que Você Saiba - LIVRO-576 Págapi-3830837100% (1)
- TradDocumento22 páginasTradMarcos Carvalho LopesAinda não há avaliações
- Do Sexo A DivindadeDocumento107 páginasDo Sexo A DivindadeCláudio Henrique Caldas Mattos100% (1)
- Protocolos Exames LaboratoriaisDocumento294 páginasProtocolos Exames LaboratoriaisDeborah NogueiraAinda não há avaliações
- Protocolos Exames LaboratoriaisDocumento294 páginasProtocolos Exames LaboratoriaisDeborah NogueiraAinda não há avaliações
- Joseph Murphy CanCOes de DeusDocumento130 páginasJoseph Murphy CanCOes de DeusAlezzinhagomes100% (3)
- Manual ATBDocumento41 páginasManual ATBenzomontresolAinda não há avaliações
- Diretrizs para o Implante de Marcapasso Cardíaco PermanenteDocumento6 páginasDiretrizs para o Implante de Marcapasso Cardíaco PermanenteenzomontresolAinda não há avaliações
- Apostila Técnica CirurgicaDocumento151 páginasApostila Técnica CirurgicaLays Alves LuchiAinda não há avaliações
- SuplementoCP Nausea Vomito Final ADocumento15 páginasSuplementoCP Nausea Vomito Final AAndre JavanAinda não há avaliações
- Drogas CardiovascularesDocumento51 páginasDrogas CardiovascularesenzomontresolAinda não há avaliações
- 8 - Estimulação Cardíaca MultisítioDocumento5 páginas8 - Estimulação Cardíaca MultisítioenzomontresolAinda não há avaliações
- 5 - Sensores Dos Marcapassos Cardíacos ArtificiaisDocumento5 páginas5 - Sensores Dos Marcapassos Cardíacos ArtificiaisenzomontresolAinda não há avaliações
- Marcapasso Cardíaco Artificial - Considerações Pré e Per-OperatóriasDocumento9 páginasMarcapasso Cardíaco Artificial - Considerações Pré e Per-OperatóriasenzomontresolAinda não há avaliações
- Eletrodo Temporário de Marca-Passo (Orientações de Uso)Documento11 páginasEletrodo Temporário de Marca-Passo (Orientações de Uso)enzomontresolAinda não há avaliações
- S Marcapasso Cardiaco PDFDocumento15 páginasS Marcapasso Cardiaco PDFAlberto FariaAinda não há avaliações
- 7 - Implante de Marcapasso Cardíaco Definitivo - Aspectos CirúrgicosDocumento11 páginas7 - Implante de Marcapasso Cardíaco Definitivo - Aspectos CirúrgicosenzomontresolAinda não há avaliações
- Zoologia Dos Bovinos - AGRVETDocumento34 páginasZoologia Dos Bovinos - AGRVETjullyAinda não há avaliações
- A Dinâmica Da EstupidezDocumento6 páginasA Dinâmica Da Estupidezandreas_noletoAinda não há avaliações
- Paleontologia em Destaque Nº 68Documento190 páginasPaleontologia em Destaque Nº 68Rodrigo Giesta FigueiredoAinda não há avaliações
- Reprodução AnimalDocumento167 páginasReprodução AnimalRennan Lins0% (1)
- Atividade 1 (1) ......... 2Documento2 páginasAtividade 1 (1) ......... 2Otávio Gonzaga TavaresAinda não há avaliações
- Avaliação Diagnostica Ciências 4°ano (1) .Docx AngelaDocumento5 páginasAvaliação Diagnostica Ciências 4°ano (1) .Docx AngelaVanuza CavalcanteAinda não há avaliações
- Curso Medicina e Manejo de RepteisDocumento63 páginasCurso Medicina e Manejo de RepteisMarombaFitness29Ainda não há avaliações
- Forma e Regime Alimentar Do Animais (Resumos)Documento47 páginasForma e Regime Alimentar Do Animais (Resumos)ceciliaguiseAinda não há avaliações
- TetrapodesDocumento73 páginasTetrapodescaiocc luisAinda não há avaliações
- Ornithorhynchus. Juntamente Com As: Mamífero Austrália. Ornithorhynchidae Equidnas Monotremados Ovíparos MonotípicaDocumento7 páginasOrnithorhynchus. Juntamente Com As: Mamífero Austrália. Ornithorhynchidae Equidnas Monotremados Ovíparos MonotípicaChronoSyAinda não há avaliações
- Av. Da 6 OS VERTEBRADOSDocumento2 páginasAv. Da 6 OS VERTEBRADOSMayris CostaAinda não há avaliações
- Mamiferos PDFDocumento31 páginasMamiferos PDFFrancisco GuedesAinda não há avaliações
- Ficha-Informativa-Animais-Vertebrados-e-Invertebrados 3º AnoDocumento3 páginasFicha-Informativa-Animais-Vertebrados-e-Invertebrados 3º Anocarpraia0% (1)
- Roteiro Aula Prática Zoologia Vertebrados 023Documento9 páginasRoteiro Aula Prática Zoologia Vertebrados 023Sophie MarucciAinda não há avaliações
- ZOOLOGIADocumento87 páginasZOOLOGIArickmedAinda não há avaliações
- Eu-Jer 10 4 1937Documento15 páginasEu-Jer 10 4 1937Lucas ArantesAinda não há avaliações
- Bestiário Aumyr 2Documento162 páginasBestiário Aumyr 2Anderson Pereira de AbreuAinda não há avaliações
- Aparelho Reprodutor 27-9-2020 PDFDocumento3 páginasAparelho Reprodutor 27-9-2020 PDFLeila Cristina AlmeidaAinda não há avaliações
- Evolucao Uma Teoria em Crise PDFDocumento4 páginasEvolucao Uma Teoria em Crise PDFJairoAinda não há avaliações
- 18 - Repteis e Aves MEDDocumento9 páginas18 - Repteis e Aves MEDAndryo CardosoAinda não há avaliações
- Ficha de Estudo Do MeioDocumento6 páginasFicha de Estudo Do MeiolinaluzAinda não há avaliações
- Infantil Iv/V: A BaleiaDocumento21 páginasInfantil Iv/V: A BaleiaLikian silvaAinda não há avaliações
- Ciencias Aparelho ReprodutorDocumento9 páginasCiencias Aparelho ReprodutorCris VieiraAinda não há avaliações
- ContagemDocumento4 páginasContagemCaio ZupaAinda não há avaliações
- Prefeitura Municipal de Bauru: Conteúdos EssenciaisDocumento47 páginasPrefeitura Municipal de Bauru: Conteúdos EssenciaisCristiane TroinaAinda não há avaliações
- Biologia 8Documento278 páginasBiologia 8Mutondo ManejaAinda não há avaliações
- Protocordados e VertebradosDocumento46 páginasProtocordados e VertebradosHigor DantasAinda não há avaliações
- Trabalho 7º AnoDocumento3 páginasTrabalho 7º AnoJéssica LayneAinda não há avaliações
- Cordados II Amniotas - SlidesDocumento36 páginasCordados II Amniotas - SlidesShizukano TsumiAinda não há avaliações
- Monografia Factores Que Influenciaram Na Determinação Do Sexo e Dimorfismo Sexual Nos RépteisDocumento26 páginasMonografia Factores Que Influenciaram Na Determinação Do Sexo e Dimorfismo Sexual Nos Répteisripanga100% (1)