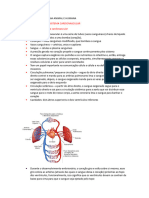
Você também pode gostar
- Apostila PilatesDocumento93 páginasApostila PilatesMatheus Camargo93% (14)
- Venuti 2021Documento354 páginasVenuti 2021Caroline PradoAinda não há avaliações
- 1 PeríodoDocumento15 páginas1 PeríodosantiagocordeirosilvaAinda não há avaliações
- O Transporte Nos Animais e FermentaçãoDocumento7 páginasO Transporte Nos Animais e FermentaçãoXavier VideiraAinda não há avaliações
- Biologia e Geologia - 10º Ano - O Transporte Nos AnimaisDocumento27 páginasBiologia e Geologia - 10º Ano - O Transporte Nos AnimaisHugo Martins100% (3)
- O Transporte Nos AnimaisDocumento9 páginasO Transporte Nos AnimaisAEESGPAinda não há avaliações
- Sistema Circulatório BiologiaDocumento9 páginasSistema Circulatório BiologiaEdmundo Cardoso CardosoAinda não há avaliações
- Aula 03 BiologiaDocumento22 páginasAula 03 BiologiaDenise NarcisoAinda não há avaliações
- PDF CONHECIMENTOS ESPECIFICOS - PROGRAMA DE CIENCIAS - Sistema CirculatórioDocumento7 páginasPDF CONHECIMENTOS ESPECIFICOS - PROGRAMA DE CIENCIAS - Sistema CirculatórioAJ JBAinda não há avaliações
- Transporte Nos Animais ResumoDocumento11 páginasTransporte Nos Animais ResumoMarcy AlexandraAinda não há avaliações
- Chadoro TrabalhoDocumento12 páginasChadoro TrabalhoCustódio Araújo MiquitaioAinda não há avaliações
- Transporte Nos AnimaisDocumento38 páginasTransporte Nos AnimaisMagda RodriguesAinda não há avaliações
- Trabalho de Fredy - BiologiaDocumento12 páginasTrabalho de Fredy - BiologiaWorld Music GroupAinda não há avaliações
- Transporte Nos AnimaisDocumento5 páginasTransporte Nos AnimaisMariana DiasAinda não há avaliações
- Transporte Nos AnimaisDocumento6 páginasTransporte Nos Animaismadalena rodriguesAinda não há avaliações
- Transporte Nos Animais PDFDocumento6 páginasTransporte Nos Animais PDFGildaMarisaAinda não há avaliações
- ##APOSTILA - Fisiologia - Sistema CARDIOVASCULAR - 2023.2Documento19 páginas##APOSTILA - Fisiologia - Sistema CARDIOVASCULAR - 2023.2marcksf77Ainda não há avaliações
- Sistema Circulatório Do Homem e Dos Animais !!!!!!!!!!!!!!!!!Documento13 páginasSistema Circulatório Do Homem e Dos Animais !!!!!!!!!!!!!!!!!Paulderdice CostaAinda não há avaliações
- Resumos - Transporte Nos AnimaisDocumento6 páginasResumos - Transporte Nos AnimaisMatilde PedroAinda não há avaliações
- Transporte Nos AnimaisDocumento28 páginasTransporte Nos AnimaisVladyslavAinda não há avaliações
- Sistema CirculatórioDocumento7 páginasSistema CirculatórioGheter CostaAinda não há avaliações
- Sistema CirculatórioDocumento17 páginasSistema CirculatórioJessica Heloisa FigueiredoAinda não há avaliações
- CIRCULATORIODocumento7 páginasCIRCULATORIOMariaEduardaAraujoAinda não há avaliações
- Fisiologia Humana E Comparada P Prof. Bruna Klassa: Sistema Cardiovascular HumanoDocumento11 páginasFisiologia Humana E Comparada P Prof. Bruna Klassa: Sistema Cardiovascular HumanoLucas GuedesAinda não há avaliações
- Biologia 5 - Sistema CardiovascularDocumento5 páginasBiologia 5 - Sistema CardiovascularRejane RedonAinda não há avaliações
- Biologia 10ºano Resumos - 1º TesteDocumento3 páginasBiologia 10ºano Resumos - 1º TesteBeatriz CortezAinda não há avaliações
- 5.2. Transporte Nos AnimaisDocumento4 páginas5.2. Transporte Nos AnimaisDiana BrancoAinda não há avaliações
- Material Completo de Apoio, Anatomia Humana II. Prof. A. Chitungo - 105719 - 113721Documento96 páginasMaterial Completo de Apoio, Anatomia Humana II. Prof. A. Chitungo - 105719 - 113721divysfinance123Ainda não há avaliações
- Sistema Circulatório - Wikipédia, A Enciclopédia LivreDocumento37 páginasSistema Circulatório - Wikipédia, A Enciclopédia LivreS4LTYAinda não há avaliações
- Sistema de TransporteDocumento3 páginasSistema de TransporteAnaAinda não há avaliações
- Conteudo P2Documento12 páginasConteudo P2fehbondezanAinda não há avaliações
- Hipótese Da Pressão RadicularDocumento3 páginasHipótese Da Pressão RadicularAna CláudiaAinda não há avaliações
- Sistema CirculatórioDocumento46 páginasSistema CirculatórioAndreaOliveira28Ainda não há avaliações
- SISTEMA CIRCULATÓRIO Aula IDocumento35 páginasSISTEMA CIRCULATÓRIO Aula IPalloma HolandaAinda não há avaliações
- Anatomia Livro-Texto - Unidade IIDocumento96 páginasAnatomia Livro-Texto - Unidade IImsspriteAinda não há avaliações
- Sistema Circulatorio - BiologiaDocumento16 páginasSistema Circulatorio - BiologiaBaptista ZaqueuAinda não há avaliações
- O Sistema CardiovascularDocumento1 páginaO Sistema CardiovascularLina MagalhaesAinda não há avaliações
- EvoluçaoDocumento6 páginasEvoluçaoJervàcio ChambalAinda não há avaliações
- Apostila Ney Av1Documento41 páginasApostila Ney Av1SuLpsAinda não há avaliações
- Aula 3Documento13 páginasAula 3Vitor PinheiroAinda não há avaliações
- Introdução - Sistema de Transporte Nos AnimaisDocumento11 páginasIntrodução - Sistema de Transporte Nos AnimaisDolinick VontadeAinda não há avaliações
- 7 Transporte Nos Animais Leya BG 10º AnoDocumento21 páginas7 Transporte Nos Animais Leya BG 10º AnoPedro B. SilvaAinda não há avaliações
- Anatomofisiologia Comparada Laboratório 2Documento8 páginasAnatomofisiologia Comparada Laboratório 2Adilson JuniorAinda não há avaliações
- BG10 - Transporte Nos AnimaisDocumento36 páginasBG10 - Transporte Nos Animaiskiko PereiraAinda não há avaliações
- Drenagem Linfática MóduloDocumento47 páginasDrenagem Linfática MóduloMagno Zaltron100% (2)
- QUESTIONÁRIO Biologia 27.11Documento4 páginasQUESTIONÁRIO Biologia 27.11Thainá MatosAinda não há avaliações
- Transporte Nos AnimaisDocumento17 páginasTransporte Nos AnimaisSara SilvaAinda não há avaliações
- Aspectos Físicos Da CirculaçãoDocumento8 páginasAspectos Físicos Da CirculaçãoCris Theodoro PoundsAinda não há avaliações
- Sistema CardiovascularDocumento7 páginasSistema CardiovascularneivaprojetosAinda não há avaliações
- Circulações Comparada e HumanaDocumento16 páginasCirculações Comparada e HumanaBreno RiveroAinda não há avaliações
- Resumo 9 - Transporte Nos AnimaisDocumento5 páginasResumo 9 - Transporte Nos AnimaisBeatriz CostaAinda não há avaliações
- Fisiologia Cardiovascular ResumoDocumento21 páginasFisiologia Cardiovascular ResumoDerek RandolphAinda não há avaliações
- 3 - Distribuição de MatériaDocumento2 páginas3 - Distribuição de MatériaFranciscoPaixão100% (1)
- Trabalho de BiologiaDocumento16 páginasTrabalho de BiologiaVictor GiembraAinda não há avaliações
- Trabalho Bio 10Documento5 páginasTrabalho Bio 10Filipe FreitasAinda não há avaliações
- Ciências Resumos Fevereiro 9anoDocumento6 páginasCiências Resumos Fevereiro 9anoMatildeAinda não há avaliações
- Capa Do Trabalho de AnatoniaDocumento15 páginasCapa Do Trabalho de AnatoniaTainan Lemos AmaralAinda não há avaliações
- O Sistema CardiovascularDocumento3 páginasO Sistema CardiovascularFrancisco GuambeAinda não há avaliações
- FISIOLOGIA Av 1Documento24 páginasFISIOLOGIA Av 1Caroline MoreiraAinda não há avaliações
- O Transporte Nos AnimaisDocumento2 páginasO Transporte Nos AnimaisAna Sanheiro100% (1)
- Benzina Retificada PDFDocumento10 páginasBenzina Retificada PDFMarcus ViniciusAinda não há avaliações
- C Naturais 6 AnoDocumento4 páginasC Naturais 6 AnodorinamendoncaAinda não há avaliações
- Tecnica PSC Processo CoachingDocumento28 páginasTecnica PSC Processo CoachingRaizes E Asas100% (1)
- 03 - Difusão Dos Gases, Transporte Dos Gases Pelo 03 - 060532Documento21 páginas03 - Difusão Dos Gases, Transporte Dos Gases Pelo 03 - 060532Ana Beatriz Moraes Da SilvaAinda não há avaliações
- 640-Rr100 Primer Skyfast Hs Cinza Rr100Documento9 páginas640-Rr100 Primer Skyfast Hs Cinza Rr100Fabrício LopesAinda não há avaliações
- Atividades 35 E 36 402 LUCIODocumento16 páginasAtividades 35 E 36 402 LUCIOmichelle lageAinda não há avaliações
- Síndrome de DownDocumento9 páginasSíndrome de DownCaio AugustoAinda não há avaliações
- Sintomas MediunidadeDocumento9 páginasSintomas MediunidadeErlania Lima de OliveiraAinda não há avaliações
- Ciencias 5 6 7 8 9Documento24 páginasCiencias 5 6 7 8 9Anonymous heAnRxNFmAinda não há avaliações
- Caderno de Canto PlatinaDocumento20 páginasCaderno de Canto PlatinaCleber AndradeAinda não há avaliações
- Aquecimento Trombone PDFDocumento42 páginasAquecimento Trombone PDFSamuel SilvaAinda não há avaliações
- Modelo de PCMSODocumento15 páginasModelo de PCMSOFábio José da SilvaAinda não há avaliações
- Sistema RespiratórioDocumento13 páginasSistema RespiratórioLUIS CLAUDIO DE BARROS PEREIRAAinda não há avaliações
- Aparelho RespiratórioDocumento14 páginasAparelho RespiratórioTC Nelinho CandidoAinda não há avaliações
- Anatomia e Fisiologia Do Sistema RespiratórioDocumento20 páginasAnatomia e Fisiologia Do Sistema Respiratórionatallia MorgannaAinda não há avaliações
- Conserva de Bananas CultivoDocumento3 páginasConserva de Bananas CultivoIfáláwoniléAgboléObemoAinda não há avaliações
- Ae ccn6 Teste 2Documento7 páginasAe ccn6 Teste 2XandraAinda não há avaliações
- Roteiro Prática Mediúnica - Rev FinalDocumento56 páginasRoteiro Prática Mediúnica - Rev FinalGiovana silva100% (3)
- Planificação 10 BG 21 - 22 V2Documento7 páginasPlanificação 10 BG 21 - 22 V2Isaura SilvaAinda não há avaliações
- Tanatologia Forense AULA 01Documento128 páginasTanatologia Forense AULA 01Douglas TostaAinda não há avaliações
- OsDocumento3 páginasOsVictor Hugo SouzaAinda não há avaliações
- Projeto o Ar Que Respiramos 3 Ano BDocumento22 páginasProjeto o Ar Que Respiramos 3 Ano BZuza, Elisangela CarvalhoAinda não há avaliações
- Ventilador Pulmonar Achieva SPO2Documento221 páginasVentilador Pulmonar Achieva SPO2Alef OliveiraAinda não há avaliações
- Resumos de Biologia 10º AnoDocumento16 páginasResumos de Biologia 10º AnoJosé M. AlmeidaAinda não há avaliações
- Sinais Vitais PediatriaDocumento5 páginasSinais Vitais PediatriaDouglas VarellaAinda não há avaliações
- 18 Exercicios de Mindfulness para CriançasDocumento9 páginas18 Exercicios de Mindfulness para CriançasEverton RobertoAinda não há avaliações
- Fisiologia e Ventilação MecânicaDocumento43 páginasFisiologia e Ventilação MecânicaDoborowskyAinda não há avaliações
- Portifolio de AnatomiaDocumento20 páginasPortifolio de AnatomiaMariana PaivaAinda não há avaliações