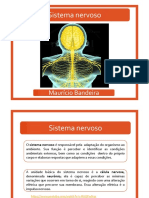
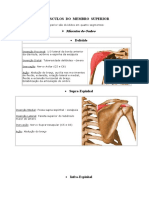
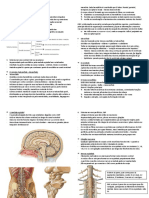
Você também pode gostar
- Aula 6-Sist - NervosoDocumento29 páginasAula 6-Sist - NervosoMárcio Stelvio Miranda da SilvaAinda não há avaliações
- Neuroanatomia: divisões e células do sistema nervosoDocumento48 páginasNeuroanatomia: divisões e células do sistema nervosoWidek Asfalio100% (1)
- Sistema NervosoDocumento45 páginasSistema NervosoduhenriqueAinda não há avaliações
- Neurofisiologia da visão e processamento de imagens no córtex visualDocumento4 páginasNeurofisiologia da visão e processamento de imagens no córtex visualandreamokadaAinda não há avaliações
- Sistema nervoso - 28 minDocumento15 páginasSistema nervoso - 28 minOssifo AliAinda não há avaliações
- Sistema Nervoso Central - CompletoDocumento182 páginasSistema Nervoso Central - CompletoProfessora Tati NaffahAinda não há avaliações
- Resumo de FisiologiaDocumento5 páginasResumo de Fisiologiajoana.rodriguesmbAinda não há avaliações
- Nervos Espinais 31Documento18 páginasNervos Espinais 31Márcio Stelvio Miranda da SilvaAinda não há avaliações
- Histologia Tecido NervosoDocumento12 páginasHistologia Tecido NervosomarianyAinda não há avaliações
- Resumo - Sistema NervosoDocumento8 páginasResumo - Sistema NervosoThayane MoreiraAinda não há avaliações
- Anatomia do Sistema Nervoso e DigestórioDocumento13 páginasAnatomia do Sistema Nervoso e DigestórioTainá CostaAinda não há avaliações
- Células Nervosas e Divisão do Sistema NervosoDocumento8 páginasCélulas Nervosas e Divisão do Sistema NervosoWesley Ramalho SouzaAinda não há avaliações
- histologiaDocumento18 páginashistologiasuyaneAinda não há avaliações
- Sistema nervosoDocumento62 páginasSistema nervosoMarianamgt100% (1)
- Aula 9. Sistema NervosoDocumento59 páginasAula 9. Sistema NervosoValdilene AndradeAinda não há avaliações
- Identificar Os Componentes Do SNP : Resumo Sistema Nervoso PeriféricoDocumento5 páginasIdentificar Os Componentes Do SNP : Resumo Sistema Nervoso PeriféricoKarolina PingarilhoAinda não há avaliações
- Aula 15 - Anatomia e Fisiologia Do Sistema Nervoso (1)Documento51 páginasAula 15 - Anatomia e Fisiologia Do Sistema Nervoso (1)Hélio SantosAinda não há avaliações
- Sistema nervoso autônomo e simpáticoDocumento10 páginasSistema nervoso autônomo e simpáticoLARISSA DE FATIMA DA SILVAAinda não há avaliações
- Introdução Ao Sistema NervosoDocumento6 páginasIntrodução Ao Sistema NervosoAnne GabrielleAinda não há avaliações
- Aula 3 SNC-SNPDocumento54 páginasAula 3 SNC-SNPBeatriz BarbosaAinda não há avaliações
- SNP II 2016 pdfDocumento33 páginasSNP II 2016 pdfAna Luiza MenezesAinda não há avaliações
- Neurofisiologia do Comportamento emDocumento11 páginasNeurofisiologia do Comportamento emFlavio CoelhoAinda não há avaliações
- Sistema Nervoso: Função e EstruturaDocumento61 páginasSistema Nervoso: Função e EstruturaBruno Avelar100% (1)
- Slide Neuro 1Documento21 páginasSlide Neuro 1Thaísa Ribeiro VieiraAinda não há avaliações
- Aula 10 - Sistema Nervoso PeriféricoDocumento34 páginasAula 10 - Sistema Nervoso PeriféricoTatiane SundstronAinda não há avaliações
- Apostila Do Sistema NervosoDocumento7 páginasApostila Do Sistema NervosoRodrigo NascimentoAinda não há avaliações
- Sistema NervosoDocumento5 páginasSistema Nervososuzanna cassarAinda não há avaliações
- Sistema Nervoso HumanoDocumento57 páginasSistema Nervoso HumanomariaAinda não há avaliações
- Resumo Sistema NervosoDocumento10 páginasResumo Sistema NervosoR7Ainda não há avaliações
- Sistema nervoso: estrutura e funçõesDocumento8 páginasSistema nervoso: estrutura e funçõesDavid JoséAinda não há avaliações
- Aula Sistema Nervoso FufsDocumento48 páginasAula Sistema Nervoso FufsJesse Galeao100% (2)
- Sistema NervosoDocumento59 páginasSistema NervosoEdu100% (1)
- Sistema Nervoso e NeurôniosDocumento29 páginasSistema Nervoso e NeurôniosLeni BarrosAinda não há avaliações
- O Sistema Nervoso CentralDocumento7 páginasO Sistema Nervoso CentralTamires AlmeidaAinda não há avaliações
- Sistema NervosoDocumento56 páginasSistema NervosoFabiana SoutoAinda não há avaliações
- Sistema NervosoDocumento41 páginasSistema NervosoJoão LucasAinda não há avaliações
- Sistema NervosoDocumento47 páginasSistema Nervosoamerica100% (1)
- Sistema nervoso: estrutura, células e funçõesDocumento65 páginasSistema nervoso: estrutura, células e funçõesWilliamAinda não há avaliações
- Sistema NervosoDocumento6 páginasSistema NervosoMiryan Mendes da SilvaAinda não há avaliações
- Sistema NervosoDocumento7 páginasSistema Nervosomarims96Ainda não há avaliações
- Sistema Nervoso PDFDocumento3 páginasSistema Nervoso PDFsuzanaAinda não há avaliações
- Sistema nervoso central e periféricoDocumento6 páginasSistema nervoso central e periféricoLetícia KlavaAinda não há avaliações
- Sistema NervosoDocumento26 páginasSistema Nervosonataliacristinedeoliveira8502Ainda não há avaliações
- Monitoria de Neuro Prova I (1) OKDocumento19 páginasMonitoria de Neuro Prova I (1) OKThaísa Ribeiro VieiraAinda não há avaliações
- Resumo Sistema NervosoDocumento4 páginasResumo Sistema NervosoCarol MoraesAinda não há avaliações
- Sistema NervosoDocumento4 páginasSistema NervosoLeonor PereiraAinda não há avaliações
- Tecido NervosoDocumento6 páginasTecido Nervosomariana machadoAinda não há avaliações
- Sistema Nervoso Central e PeriféricoDocumento3 páginasSistema Nervoso Central e PeriféricoYthisack Silva50% (2)
- APOSTILA DO SISTEMA NERVOSODocumento9 páginasAPOSTILA DO SISTEMA NERVOSOSheila LacerdaAinda não há avaliações
- O cérebro e o sistema nervoso: a estrutura e as funçõesDocumento5 páginasO cérebro e o sistema nervoso: a estrutura e as funçõesSara JesusAinda não há avaliações
- Sistem Nervoso 4. ExercíciosDocumento4 páginasSistem Nervoso 4. ExercíciosAna AntiqueiraAinda não há avaliações
- S.nervoso NeuroniosDocumento31 páginasS.nervoso NeuroniosPauloArmandoPedroJonasAinda não há avaliações
- Aula 2Documento49 páginasAula 2Leticia MaggioriniAinda não há avaliações
- Sistema nervoso: conceito, estrutura e funçõesDocumento45 páginasSistema nervoso: conceito, estrutura e funçõesBruna TelisAinda não há avaliações
- Relatório Problema 2 - ASE 4Documento12 páginasRelatório Problema 2 - ASE 4felipeborges39949Ainda não há avaliações
- Sistema nervoso autônomo e somático: anatomia e funções geraisDocumento4 páginasSistema nervoso autônomo e somático: anatomia e funções geraisAnne MartiniAinda não há avaliações
- Sistema Nervoso emDocumento47 páginasSistema Nervoso emEnzo AlegreAinda não há avaliações
- SNC vs SNP e divisões anatômica e funcional do SNDocumento8 páginasSNC vs SNP e divisões anatômica e funcional do SNAriel Gomes100% (1)
- NeurociênciasDocumento6 páginasNeurociênciasAmanda YaniAinda não há avaliações
- Temática - Ciências da Vida - Relação e CoordenaçãoNo EverandTemática - Ciências da Vida - Relação e CoordenaçãoAinda não há avaliações
- Resumo Sistema Nervoso AutônomoDocumento3 páginasResumo Sistema Nervoso AutônomotamiossopolianaAinda não há avaliações
- Cerebelo - Morfologia e FunçõesDocumento2 páginasCerebelo - Morfologia e FunçõesJediel PedrosoAinda não há avaliações
- Sistema nervoso e suas funçõesDocumento105 páginasSistema nervoso e suas funçõesKelly AlvesAinda não há avaliações
- 2 - Princípios Fundamentais em NeurologiaDocumento45 páginas2 - Princípios Fundamentais em NeurologiaCaíque BarrosAinda não há avaliações
- Fisio SistemasDocumento69 páginasFisio Sistemaskarine.inacio50% (1)
- Músculos do ombro e membro superiorDocumento7 páginasMúsculos do ombro e membro superiorMarcelo Cruz de Oliveira100% (1)
- Síndromes MedularesDocumento38 páginasSíndromes MedularesCriz LuzAinda não há avaliações
- Neuroanatomia - exercício sobre estruturas da medula e nervos cranianosDocumento4 páginasNeuroanatomia - exercício sobre estruturas da medula e nervos cranianosLuiz EduardoAinda não há avaliações
- Função motora e distúrbiosDocumento17 páginasFunção motora e distúrbiosSara CastroAinda não há avaliações
- Revista DOR Complexo Trigeminal e DepressaoDocumento32 páginasRevista DOR Complexo Trigeminal e DepressaoKATIANE SMANIOTTO TRESAinda não há avaliações
- Sistema nervoso autônomo e simpáticoDocumento10 páginasSistema nervoso autônomo e simpáticoLARISSA DE FATIMA DA SILVAAinda não há avaliações
- Anatomia da medula espinhal emDocumento5 páginasAnatomia da medula espinhal emEDSONAinda não há avaliações
- Introdução Ao Sistema Nervoso 2022Documento21 páginasIntrodução Ao Sistema Nervoso 2022Kathleen Saraiva0% (1)
- Aula 2 - Casos Cli - Nicos Com Paciente Com Problemas Na ColunaDocumento13 páginasAula 2 - Casos Cli - Nicos Com Paciente Com Problemas Na ColunaFranciscoAinda não há avaliações
- Aula 9 - Medula EspinhalDocumento80 páginasAula 9 - Medula EspinhalAlessandra OliveiraAinda não há avaliações
- Plano de Curso NeuroanatomiaDocumento3 páginasPlano de Curso NeuroanatomiaArnailton A. BentoAinda não há avaliações
- Aula 01 - Sistema NervosoDocumento7 páginasAula 01 - Sistema NervosoSabrina SilvaAinda não há avaliações
- Anatomia Dos Nervos CranianosDocumento5 páginasAnatomia Dos Nervos CranianosGlória Mamédio100% (1)
- Nervos CranianosDocumento7 páginasNervos CranianosCarla_Cristina_6798Ainda não há avaliações
- Sistema Nervoso Visceral e suas FunçõesDocumento15 páginasSistema Nervoso Visceral e suas FunçõesAlexandre SobralAinda não há avaliações
- O Sistema Nervoso e suas partesDocumento2 páginasO Sistema Nervoso e suas partesGabriel RibeiroAinda não há avaliações
- Anatomia Macroscopica Da Medula EspinhalDocumento4 páginasAnatomia Macroscopica Da Medula EspinhalKimã Barbosa MeiraAinda não há avaliações
- Atividade Anatomia Sistema NervosoDocumento3 páginasAtividade Anatomia Sistema NervosoAna Clara R. LanaAinda não há avaliações
- Sistema activador reticular ascendente (SARA) y su función en la regulación del sueño y la vigiliaDocumento2 páginasSistema activador reticular ascendente (SARA) y su función en la regulación del sueño y la vigiliaChristianAnagorAinda não há avaliações
- Sistema NervosoDocumento4 páginasSistema NervosoLeonor PereiraAinda não há avaliações
- Ficha de Leitura - SNDocumento3 páginasFicha de Leitura - SNsaidefelix mariaAinda não há avaliações
- Neuroanatomia emDocumento6 páginasNeuroanatomia emobjetivoibi carvalhoAinda não há avaliações
- Controle Encefálico Do MovimentoDocumento4 páginasControle Encefálico Do MovimentoFisiologia IAinda não há avaliações
- Lesoes Do Plexo BraquialDocumento5 páginasLesoes Do Plexo BraquialJanayna AndradeAinda não há avaliações