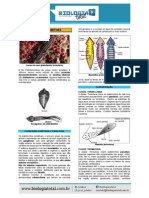
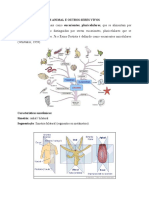
Você também pode gostar
- PlatelmintosDocumento1 páginaPlatelmintosBianca Gabrielly Oliveira SilvaAinda não há avaliações
- PlatelmintosDocumento22 páginasPlatelmintosisabel AndradeAinda não há avaliações
- Platelmintos e NematódeosDocumento10 páginasPlatelmintos e NematódeosDailton NhantumboAinda não há avaliações
- É Extremamente Importante para o Entendimento Da Genética CompreenderDocumento11 páginasÉ Extremamente Importante para o Entendimento Da Genética CompreenderPatrício AntónioAinda não há avaliações
- Reino Animal VermesDocumento15 páginasReino Animal VermesgustavolancinikrempelAinda não há avaliações
- Os principais vermes parasitas e de vida livre: PlatelmintesDocumento6 páginasOs principais vermes parasitas e de vida livre: Platelmintesmariaveronica1Ainda não há avaliações
- Platelmintos e Nematódeos: Características e DoençasDocumento1 páginaPlatelmintos e Nematódeos: Características e DoençasBruno TakahashiAinda não há avaliações
- platelmintos- resumo e exercíciosDocumento2 páginasplatelmintos- resumo e exercíciosBRUNA GABRIELA BOGORNIAinda não há avaliações
- Platelmintos e NematelmintosDocumento13 páginasPlatelmintos e NematelmintosHannah LuísaAinda não há avaliações
- Os Platelmintos: André MaiaDocumento5 páginasOs Platelmintos: André MaiaDailton NhantumboAinda não há avaliações
- Reino Animal - Poríferos, Cnidários e PlatelmintosDocumento21 páginasReino Animal - Poríferos, Cnidários e PlatelmintosBruno FelipeAinda não há avaliações
- Bacteiras e Seus PerigosDocumento2 páginasBacteiras e Seus PerigosJose SilvasAinda não há avaliações
- Filo Platyhelminthes e NematyhelminthesDocumento11 páginasFilo Platyhelminthes e NematyhelminthesMateusMarinhoAinda não há avaliações
- Universidade Federal do Maranhão ensina sobre o filo NematodaDocumento8 páginasUniversidade Federal do Maranhão ensina sobre o filo NematodaHyngrid HevilynAinda não há avaliações
- Trabalho BiologiaDocumento17 páginasTrabalho Biologiadelimaanaclara1Ainda não há avaliações
- Platelmitos e Nematelmintos TabelaDocumento3 páginasPlatelmitos e Nematelmintos TabelamarianaAinda não há avaliações
- Reino Animal - Invertebrados e VermesDocumento234 páginasReino Animal - Invertebrados e VermesCaroline Angeli SancioAinda não há avaliações
- Zoologia 01Documento2 páginasZoologia 01samuel3portelaAinda não há avaliações
- 5 HelmintosDocumento28 páginas5 HelmintosDenivaldo Silva FerreiraAinda não há avaliações
- Patologia e Parasitologia 8Documento16 páginasPatologia e Parasitologia 8Adilson JuniorAinda não há avaliações
- Características dos PlatelmintosDocumento7 páginasCaracterísticas dos PlatelmintosAry JuniorAinda não há avaliações
- Platelmintas: Características e ClassesDocumento6 páginasPlatelmintas: Características e ClassesCarolina Enxuga Sá MartinsAinda não há avaliações
- Platelmintos: vermes de corpo achatadoDocumento12 páginasPlatelmintos: vermes de corpo achatadoSamuel XzwAinda não há avaliações
- HelmintosDocumento5 páginasHelmintosluisAinda não há avaliações
- Reino Animal: Características dos principais filosDocumento11 páginasReino Animal: Características dos principais filosEzimira SaulAinda não há avaliações
- Adaptações de animais enterrados e respiração pulmonarDocumento39 páginasAdaptações de animais enterrados e respiração pulmonarKenzel Kache VerborgeAinda não há avaliações
- Reino Animal - 3º AnoDocumento5 páginasReino Animal - 3º Anokimtete290296Ainda não há avaliações
- Reino Animalia - Prof. AnacletoDocumento40 páginasReino Animalia - Prof. AnacletoHiper JuniorAinda não há avaliações
- Reinos animais e seus folhetos embrionáriosDocumento12 páginasReinos animais e seus folhetos embrionáriosNina PolitanoAinda não há avaliações
- Platelmintos 3 OanoemDocumento30 páginasPlatelmintos 3 OanoemLUCIANAAinda não há avaliações
- Animais AncelomadosDocumento15 páginasAnimais AncelomadosHelton Victor Camizola CamizolaAinda não há avaliações
- IntroduçãoDocumento21 páginasIntroduçãoMiguel Manuel JoãoAinda não há avaliações
- Apostila de Zoologia GeralDocumento30 páginasApostila de Zoologia GeralJulia MaiaAinda não há avaliações
- PLATELMINTOSDocumento9 páginasPLATELMINTOSGustavo de BarrosAinda não há avaliações
- Diferenças entre animais e outros seres vivosDocumento17 páginasDiferenças entre animais e outros seres vivosEntraro NobaroAinda não há avaliações
- Platelmintos: vermes achatados parasitas do homemDocumento10 páginasPlatelmintos: vermes achatados parasitas do homemThiago RibeiroAinda não há avaliações
- Giad 03Documento45 páginasGiad 03EDUARDO CERQUEIRA E SILVA100% (1)
- Relatório sobre os principais vermes parasitasDocumento4 páginasRelatório sobre os principais vermes parasitasbarbaraverenaAinda não há avaliações
- Zoologia de invertebrados: características e filosDocumento27 páginasZoologia de invertebrados: características e filosÂngela DiasAinda não há avaliações
- Eja 19-10Documento6 páginasEja 19-10ELLEN CAMILE DA SILVAAinda não há avaliações
- Reino Animalia: Características GeraisDocumento6 páginasReino Animalia: Características GeraisMauro Miguel MartinsAinda não há avaliações
- Animalia – Wikipédia, A Enciclopédia LivreDocumento40 páginasAnimalia – Wikipédia, A Enciclopédia LivreKelvin Fernando NovidadesAinda não há avaliações
- Celenterados e PlatelmintosDocumento1 páginaCelenterados e PlatelmintosDiego PaesAinda não há avaliações
- Introdução aos invertebradosDocumento4 páginasIntrodução aos invertebradosLUCAS FONSECA DE OLIVEIRAAinda não há avaliações
- Características gerais dos poríferos, cnidários, platelmintos e nematódeosDocumento107 páginasCaracterísticas gerais dos poríferos, cnidários, platelmintos e nematódeosdibugiu100% (1)
- Apostila de Zoologia-BDocumento61 páginasApostila de Zoologia-BscaaaaAinda não há avaliações
- Filo NematelmintosDocumento11 páginasFilo NematelmintosMiguel Da PlacaAinda não há avaliações
- Zoologia Dos Invertebrados e Vertebrados: Pré-Vestibular Dimensão, Biologia, Profa Roberta TroisDocumento33 páginasZoologia Dos Invertebrados e Vertebrados: Pré-Vestibular Dimensão, Biologia, Profa Roberta TroisAntonio Fernando LimaAinda não há avaliações
- Reino Animalia - Evolução e CaracterísticasDocumento5 páginasReino Animalia - Evolução e CaracterísticasInfospace100% (1)
- Os principais platelmintos e nematódeos parasitas humanosDocumento4 páginasOs principais platelmintos e nematódeos parasitas humanoslaters codfeeAinda não há avaliações
- Platelmintos adaptaçõesDocumento22 páginasPlatelmintos adaptaçõesAldemir Gabriel100% (1)
- Guia de Sobrevivência - ZoologiaDocumento10 páginasGuia de Sobrevivência - Zoologiaana luizaAinda não há avaliações
- Parasitoses IntestinalDocumento2 páginasParasitoses IntestinaleugeniotgomesAinda não há avaliações
- ProtozoáriosDocumento42 páginasProtozoáriosHaris HarahapAinda não há avaliações
- Trabalho de Biologia A - 1º Bimestre (Módulo 1,2,3 e 4)Documento21 páginasTrabalho de Biologia A - 1º Bimestre (Módulo 1,2,3 e 4)Sandra CardosoAinda não há avaliações
- Zoologia de InvertebradosDocumento9 páginasZoologia de InvertebradosSuelen GomesAinda não há avaliações
- Relatório de Invertebrados ZoologiaDocumento4 páginasRelatório de Invertebrados ZoologiajessicaAinda não há avaliações
- TCC JoaoAltenfelderFinalDocumento80 páginasTCC JoaoAltenfelderFinalHerminio JuvenalsAinda não há avaliações
- Monografia - Alice Tavares de OliveiraDocumento63 páginasMonografia - Alice Tavares de OliveiraHerminio JuvenalsAinda não há avaliações
- Aula 6 (Unidade 3) - Sistemas de CusteioDocumento7 páginasAula 6 (Unidade 3) - Sistemas de CusteioHerminio JuvenalsAinda não há avaliações
- Gestão financeira: cálculo de indicadores de atividade, rentabilidade e liquidezDocumento3 páginasGestão financeira: cálculo de indicadores de atividade, rentabilidade e liquidezHerminio JuvenalsAinda não há avaliações
- Divisão de Economia e Gestão 202043Documento18 páginasDivisão de Economia e Gestão 202043Herminio JuvenalsAinda não há avaliações
- Casos Práticos ComercialDocumento11 páginasCasos Práticos ComercialManuel Rodrigues ImbiAinda não há avaliações
- Distribuição de Temas FP 2021Documento1 páginaDistribuição de Temas FP 2021Herminio JuvenalsAinda não há avaliações
- Finanças púb-WPS Office-1Documento8 páginasFinanças púb-WPS Office-1Herminio JuvenalsAinda não há avaliações
- Exercicios ResolvidosDocumento9 páginasExercicios ResolvidosHerminio JuvenalsAinda não há avaliações
- Curso de Contabilidade e Auditoria e Administração PúblicaDocumento2 páginasCurso de Contabilidade e Auditoria e Administração PúblicaHerminio JuvenalsAinda não há avaliações
- 2 º Teste de Contabilidade de Gestão III - DiurnoDocumento3 páginas2 º Teste de Contabilidade de Gestão III - DiurnoHerminio Juvenals100% (1)
- Avaliação de investimentos com fluxo de caixa descontadoDocumento2 páginasAvaliação de investimentos com fluxo de caixa descontadoHerminio JuvenalsAinda não há avaliações
- Documento de ArrendamentoDocumento11 páginasDocumento de ArrendamentoDaniele Gonçalves MagalhãesAinda não há avaliações
- Administração Pública: Conceitos e FinsDocumento4 páginasAdministração Pública: Conceitos e FinsHerminio JuvenalsAinda não há avaliações
- Métodos de repartição de custos indiretos ABC e secções homogéneasDocumento6 páginasMétodos de repartição de custos indiretos ABC e secções homogéneasHerminio Juvenals100% (2)
- Ficha 4 - Fundamentos Do Controlo Financeiro Da Administração PúblicaDocumento12 páginasFicha 4 - Fundamentos Do Controlo Financeiro Da Administração PúblicaHerminio JuvenalsAinda não há avaliações
- Exemplo Metodo ABCDocumento2 páginasExemplo Metodo ABCHerminio JuvenalsAinda não há avaliações
- Resumo Dos Trabalhos222Documento7 páginasResumo Dos Trabalhos222Herminio JuvenalsAinda não há avaliações
- UNIDADE II - SISTEMAS DE CUSTEIO (Revisao)Documento5 páginasUNIDADE II - SISTEMAS DE CUSTEIO (Revisao)Herminio JuvenalsAinda não há avaliações
- Sistema Fiscal MocambicanoDocumento9 páginasSistema Fiscal MocambicanoHerminio JuvenalsAinda não há avaliações
- Ficha 4 - Fundamentos Do Controlo Financeiro Da Administração PúblicaDocumento12 páginasFicha 4 - Fundamentos Do Controlo Financeiro Da Administração PúblicaHerminio JuvenalsAinda não há avaliações
- Financas Publicas Introd 2edDocumento2 páginasFinancas Publicas Introd 2edHerminio JuvenalsAinda não há avaliações
- Nir 300Documento4 páginasNir 300Herminio JuvenalsAinda não há avaliações
- Intervenção Pública Na EconomiaDocumento9 páginasIntervenção Pública Na EconomiaHerminio JuvenalsAinda não há avaliações
- Controlo Interno AuditoriaDocumento10 páginasControlo Interno AuditoriaHerminio JuvenalsAinda não há avaliações
- ISSAI - 100 - Principios Fundamentais de Auditoria Do Setor PublicoDocumento17 páginasISSAI - 100 - Principios Fundamentais de Auditoria Do Setor PublicoCeciliaMartinsAinda não há avaliações
- Ficha 1 NOCOES DE ADMINISTRAÇÃO PÚBLICA-2Documento6 páginasFicha 1 NOCOES DE ADMINISTRAÇÃO PÚBLICA-2Herminio Juvenals100% (1)
- Programa de Auditoria Financeira As Contas Das Instituições PúblicasDocumento5 páginasPrograma de Auditoria Financeira As Contas Das Instituições PúblicasHerminio JuvenalsAinda não há avaliações
- Ficha 4 - Fundamentos Do Controlo Financeiro Da Administração PúblicaDocumento12 páginasFicha 4 - Fundamentos Do Controlo Financeiro Da Administração PúblicaHerminio JuvenalsAinda não há avaliações
- ISSAI 400 PortuguesDocumento22 páginasISSAI 400 PortuguesHerminio JuvenalsAinda não há avaliações
- Ap3 Pesquisa Mercadologica Uninta - Passei DiretoDocumento11 páginasAp3 Pesquisa Mercadologica Uninta - Passei DiretoFelipe LimaAinda não há avaliações
- Diagnóstico dos 5 Ps da estratégia em empresa de usinagemDocumento3 páginasDiagnóstico dos 5 Ps da estratégia em empresa de usinagemJoão Paulo G. SilveiraAinda não há avaliações
- CatalogoDocumento72 páginasCatalogoQue torcida é essa.Ainda não há avaliações
- Habilidades e ProfissõesDocumento10 páginasHabilidades e Profissõeslucaspt1000Ainda não há avaliações
- Conceitualização Cognitiva: Pensamentos, Crenças e EstratégiasDocumento5 páginasConceitualização Cognitiva: Pensamentos, Crenças e EstratégiasMauricio Marane0% (1)
- Fibra Tipo 3Documento5 páginasFibra Tipo 3Edimilson LanaAinda não há avaliações
- Estatuto do servidor DiademaDocumento46 páginasEstatuto do servidor Diademajmaj jmajAinda não há avaliações
- Procedimentos para ensaio de viga de concreto protendidaDocumento66 páginasProcedimentos para ensaio de viga de concreto protendidaLucasAinda não há avaliações
- SimposioANPEPP 2008Documento178 páginasSimposioANPEPP 2008Hábilis Augusto BiazottoAinda não há avaliações
- Análise de Risco para Supressão VegetalDocumento12 páginasAnálise de Risco para Supressão VegetalAdilson Sousa LimaAinda não há avaliações
- A AMIZADE É MAGICA MODULO OP VFINAL - Por Onze#6904Documento27 páginasA AMIZADE É MAGICA MODULO OP VFINAL - Por Onze#6904Thecla CoronataAinda não há avaliações
- Orgânica3 Hidrocarbonetos (Parte1)Documento21 páginasOrgânica3 Hidrocarbonetos (Parte1)Lorenzo RamosAinda não há avaliações
- Comunidades de Prática Na Educação Superior A DistânciaDocumento4 páginasComunidades de Prática Na Educação Superior A DistânciakehrleAinda não há avaliações
- Procedimento Operacional Padrão: TituloDocumento7 páginasProcedimento Operacional Padrão: TituloDeyvid OliveiraAinda não há avaliações
- Halliday 3 - Cap 31Documento8 páginasHalliday 3 - Cap 31Peter DutaAinda não há avaliações
- 2815-lms FileDocumento5 páginas2815-lms FileShayrula AliceAinda não há avaliações
- Relatório Financeiro 2022Documento3 páginasRelatório Financeiro 2022Diogo FranciscoAinda não há avaliações
- A Vida de Uma Princesa Árabe Revela a Opressão das MulheresDocumento144 páginasA Vida de Uma Princesa Árabe Revela a Opressão das MulheresMarcos Bezerra da CostaAinda não há avaliações
- LIVRO DE MAT - ANDRINI - 7 Ano-2Documento1 páginaLIVRO DE MAT - ANDRINI - 7 Ano-2Ronaldo SidneiAinda não há avaliações
- Criar ConteúdoDocumento62 páginasCriar ConteúdoDavidCamarinhaAinda não há avaliações
- Física e Química - Movimento circular e retilíneoDocumento2 páginasFísica e Química - Movimento circular e retilíneoDiogo SIlvaAinda não há avaliações
- Inquerito - PFDocumento10 páginasInquerito - PFliscappiAinda não há avaliações
- Virtual Wi Fi Router Download BaixakiDocumento8 páginasVirtual Wi Fi Router Download BaixakiPauloAinda não há avaliações
- Avaliação em Educação Física AdaptadaDocumento4 páginasAvaliação em Educação Física AdaptadaBianca MeloAinda não há avaliações
- Sistema tratamento efluentes residência 4 pessoasDocumento3 páginasSistema tratamento efluentes residência 4 pessoasweber100% (1)
- Bpo - 2291-2291-1-PBDocumento18 páginasBpo - 2291-2291-1-PBalesorianoAinda não há avaliações
- Plano de Educação Ambiental para Trabalhadores - Rev01Documento7 páginasPlano de Educação Ambiental para Trabalhadores - Rev01smsengemon jequieAinda não há avaliações
- Mapa - Far - Farmacologia Aplicada - 52-2023Documento2 páginasMapa - Far - Farmacologia Aplicada - 52-2023Cavalini Assessoria AcadêmicaAinda não há avaliações
- Lista de verificação para inclusão de arma por compra na indústria nacionalDocumento1 páginaLista de verificação para inclusão de arma por compra na indústria nacionalNet WalkerAinda não há avaliações
- Modulo 2 FTFDocumento16 páginasModulo 2 FTFAmanda SáAinda não há avaliações