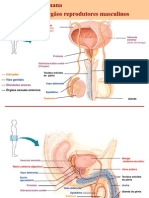
Você também pode gostar
- Cartilha Inseminacao Artificial Suinos 000g0tsklpp02wx5ok026zxpghnbhzctDocumento11 páginasCartilha Inseminacao Artificial Suinos 000g0tsklpp02wx5ok026zxpghnbhzctNatália NmsAinda não há avaliações
- Weber Max Economia e Sociedade Volume 1pdfDocumento72 páginasWeber Max Economia e Sociedade Volume 1pdfJussara Ferreira50% (2)
- 1-Coleta e Avaliao de Smen 2Documento47 páginas1-Coleta e Avaliao de Smen 2Lucas FonsecaAinda não há avaliações
- 02 Diagnostico Micologico RedDocumento58 páginas02 Diagnostico Micologico RedNatália NmsAinda não há avaliações
- Cap 24 Historia Da Riqueza Do Homem Marcia GuerraDocumento12 páginasCap 24 Historia Da Riqueza Do Homem Marcia GuerraNatália NmsAinda não há avaliações
- Weber - Tipos de DominaçãoDocumento50 páginasWeber - Tipos de DominaçãoAndré MagnelliAinda não há avaliações
- Anatomia DigestivaDocumento12 páginasAnatomia DigestivaGabriel GomesAinda não há avaliações
- Administrativo Mat CresDocumento56 páginasAdministrativo Mat CresNatália NmsAinda não há avaliações
- Deep Vol. 4 - Kylie Scott (Revisado)Documento317 páginasDeep Vol. 4 - Kylie Scott (Revisado)Natália NmsAinda não há avaliações
- 3o Disciplina - Introducao A EconomiaDocumento128 páginas3o Disciplina - Introducao A Economiasandro Mayhem100% (2)
- Freire - Consideraã Ã Es em Torno Do Ato de EstudarDocumento3 páginasFreire - Consideraã Ã Es em Torno Do Ato de EstudarNatália NmsAinda não há avaliações
- Cap 23 Historia Da Riqueza Do Homem Marcia GuerraDocumento12 páginasCap 23 Historia Da Riqueza Do Homem Marcia GuerraNatália NmsAinda não há avaliações
- ElisaDocumento2 páginasElisagordon_bayAinda não há avaliações
- Manual ImunoDocumento36 páginasManual ImunoNatália NmsAinda não há avaliações
- Arruda Et Al 2011 Revisao SemenDocumento7 páginasArruda Et Al 2011 Revisao SemenNatália NmsAinda não há avaliações
- Fundamentos de Terapêutica VeterináriaDocumento222 páginasFundamentos de Terapêutica VeterináriaAdemir Tancini0% (1)
- 270 1266 1 PBDocumento8 páginas270 1266 1 PBNatália NmsAinda não há avaliações
- Endocrinologia Da Fêmea CaninaDocumento51 páginasEndocrinologia Da Fêmea CaninaNatália NmsAinda não há avaliações
- Araujo AP Me BotfmvzDocumento139 páginasAraujo AP Me BotfmvzNatália NmsAinda não há avaliações
- Arquivo 03Documento4 páginasArquivo 03Natália NmsAinda não há avaliações
- 1126 20299 1 PBDocumento7 páginas1126 20299 1 PBNatália NmsAinda não há avaliações
- CamarãoDocumento17 páginasCamarãoNatália NmsAinda não há avaliações
- 925Documento8 páginas925Natália NmsAinda não há avaliações
- Avaliacao de Riscos Cardiacos para Procedimentos Anestesicos e CirurgicosDocumento17 páginasAvaliacao de Riscos Cardiacos para Procedimentos Anestesicos e CirurgicosJeferson Silva CarvalhoAinda não há avaliações
- A Importância Da Brucelose Bovina Na Saúde PúblicaDocumento6 páginasA Importância Da Brucelose Bovina Na Saúde PúblicaNatália NmsAinda não há avaliações
- Qualidade Do Leite CcsDocumento12 páginasQualidade Do Leite CcsDenis SatoAinda não há avaliações
- Toxoplasmose Animal No BrasilDocumento23 páginasToxoplasmose Animal No BrasilNatália NmsAinda não há avaliações
- SIMP - Assepsia e AntissepsiaDocumento9 páginasSIMP - Assepsia e AntissepsiaanaclaraferreiraAinda não há avaliações
- Candidose Na Medicina Veterinária Um Enfoque Micológico, Clínico e TerapêuticoDocumento10 páginasCandidose Na Medicina Veterinária Um Enfoque Micológico, Clínico e TerapêuticoNatália NmsAinda não há avaliações
- Guerra Da Secessao - Farid AmeurDocumento69 páginasGuerra Da Secessao - Farid AmeurNatália NmsAinda não há avaliações
- Aula 1 Resumo Ciclo MenstrualDocumento6 páginasAula 1 Resumo Ciclo MenstrualisabelAinda não há avaliações
- Relatório - Observação de Cortes Histológicos de Ovários e de Testículos em Preparações DefinitivasDocumento30 páginasRelatório - Observação de Cortes Histológicos de Ovários e de Testículos em Preparações DefinitivasMario Nascimento33% (3)
- Foca Na Medicina Questões de Biologia Professor José DrummondDocumento10 páginasFoca Na Medicina Questões de Biologia Professor José DrummondJohn JesusAinda não há avaliações
- Embriologia Ensino Superior Resumo PDFDocumento41 páginasEmbriologia Ensino Superior Resumo PDFAry Junior100% (1)
- Livro Unico PDFDocumento212 páginasLivro Unico PDFSamira FerreiraAinda não há avaliações
- IATF em Bovinos JRDocumento18 páginasIATF em Bovinos JRRayan CardosoAinda não há avaliações
- PSU Prova Geral Tipo 3 2014Documento36 páginasPSU Prova Geral Tipo 3 2014Jaime XavierAinda não há avaliações
- Fisiologia - Crescimento e DesenvolvimentoDocumento18 páginasFisiologia - Crescimento e DesenvolvimentoMariana VerlyAinda não há avaliações
- E-Book de Apoio - Saúde Hormonal e A Interação Corpo e MenteDocumento17 páginasE-Book de Apoio - Saúde Hormonal e A Interação Corpo e MenteMylena karolyne Souza silvaAinda não há avaliações
- Endocrinologia Da Reprodução de Peixes.Documento7 páginasEndocrinologia Da Reprodução de Peixes.lucileine assumpcaoAinda não há avaliações
- Biologia - 12º AnoDocumento3 páginasBiologia - 12º AnoJoão FerreiraAinda não há avaliações
- Aula 4 - Fisiologia IIDocumento9 páginasAula 4 - Fisiologia IIDeborah AmbreAinda não há avaliações
- Anticoncepcionais Hormonais Mari QuintelaDocumento8 páginasAnticoncepcionais Hormonais Mari QuintelaAfiliado LendárioAinda não há avaliações
- GINECOLOGIADocumento41 páginasGINECOLOGIAMilena CristinaAinda não há avaliações
- 4 - Regulação Hormonal Feminina e MasculinaDocumento28 páginas4 - Regulação Hormonal Feminina e MasculinaMilene100% (1)
- Aula 12 - HipopituitarismoDocumento24 páginasAula 12 - Hipopituitarismosanthiagoschneider100% (2)
- Manual IATF - BovDocumento13 páginasManual IATF - BovVitoria MaltaAinda não há avaliações
- O Sagrado FemininoDocumento20 páginasO Sagrado FemininoJaqueline Fernandes89% (9)
- Neuroendocrinologia Da ReproduçãoDocumento4 páginasNeuroendocrinologia Da ReproduçãoPablo AmadorAinda não há avaliações
- Fitoterapia e TPMDocumento133 páginasFitoterapia e TPMHelena100% (1)
- Transf Embriões Bovinos PDFDocumento20 páginasTransf Embriões Bovinos PDFJeds Sol GiovannaAinda não há avaliações
- APG 24 - Sist. Genital FemininoDocumento18 páginasAPG 24 - Sist. Genital FemininoVinicius DiasAinda não há avaliações
- Fisiologia Do Ciclo Menstrual - FinalDocumento13 páginasFisiologia Do Ciclo Menstrual - FinalFamília Cleber Celi NathanaelAinda não há avaliações
- Aspectos Nutricionais Relacionados Ao Ciclo MenstrualDocumento9 páginasAspectos Nutricionais Relacionados Ao Ciclo MenstrualGislaine LealAinda não há avaliações
- Ficha de Trabalho 2 Reproducao Humana2Documento6 páginasFicha de Trabalho 2 Reproducao Humana2Ana Canastra100% (1)
- Sistema ReprodutorDocumento11 páginasSistema ReprodutorBeaAinda não há avaliações
- Lista de Labs PDFDocumento17 páginasLista de Labs PDFAndre Jardim DominguesAinda não há avaliações
- Exercícios Sobre Sistema Hormonal Com GabaritoDocumento2 páginasExercícios Sobre Sistema Hormonal Com GabaritoSérgio CarlosAinda não há avaliações
- Ficha de Trabalho 3 Biologia 12Documento2 páginasFicha de Trabalho 3 Biologia 12Renato Costa80% (5)
- Erros Mais RecentesDocumento7 páginasErros Mais RecentesMariana NolascoAinda não há avaliações