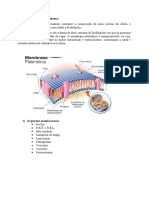
Você também pode gostar
- Expressões IdiomáticasDocumento2 páginasExpressões IdiomáticasCristian Pérez Lecaros100% (5)
- Curativos para feridas limpas e infectadasDocumento5 páginasCurativos para feridas limpas e infectadasRenata de CarvalhoAinda não há avaliações
- SNA Aula Sistema Nervoso AutônomoDocumento2 páginasSNA Aula Sistema Nervoso AutônomoSylvana NoronhaAinda não há avaliações
- Glândulas Anexas de SuínosDocumento2 páginasGlândulas Anexas de SuínosCarlos Miguel BatistaAinda não há avaliações
- Fechamento Problema 2Documento4 páginasFechamento Problema 2anna luisaAinda não há avaliações
- Membrana Celular: Transporte e Modelo Mosaico-FluidoDocumento8 páginasMembrana Celular: Transporte e Modelo Mosaico-FluidoMatilde PedroAinda não há avaliações
- Membrana Celular: Transporte e Estrutura emDocumento46 páginasMembrana Celular: Transporte e Estrutura emTatiane ValeAinda não há avaliações
- Obtenção de Matéria PDFDocumento15 páginasObtenção de Matéria PDFLeticia RussoAinda não há avaliações
- Resumo Biologia CelularDocumento27 páginasResumo Biologia CelularSalvar Meus JoguinhosAinda não há avaliações
- Estrutura e funções da membrana celularDocumento10 páginasEstrutura e funções da membrana celularJulia AraujoAinda não há avaliações
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Documento13 páginasModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixAinda não há avaliações
- Aula 2 Biofísica 24Documento22 páginasAula 2 Biofísica 24Sílvia ScabinAinda não há avaliações
- Apostila-Citologia-pg 19 A 34Documento16 páginasApostila-Citologia-pg 19 A 34denisetg100% (2)
- Biologia Celular TransporteDocumento51 páginasBiologia Celular TransporteBeatriz SPAinda não há avaliações
- Membrana CelularDocumento10 páginasMembrana CelularEsmeralda MuchisseAinda não há avaliações
- 2986-2 - Citologia - Membrana PlasmáticaDocumento6 páginas2986-2 - Citologia - Membrana PlasmáticaRênisson KalinoAinda não há avaliações
- Membranas CelularesDocumento22 páginasMembranas Celularesluisa diasAinda não há avaliações
- Obtenção de Matéria (Resumo Online)Documento7 páginasObtenção de Matéria (Resumo Online)Maria Abrantes TavaresAinda não há avaliações
- Estrutura e função das membranas celulares e sistemas de transporteDocumento14 páginasEstrutura e função das membranas celulares e sistemas de transporteLuis RicardoAinda não há avaliações
- Biologia Transporte Na CélulaDocumento3 páginasBiologia Transporte Na CélulaMárcio Simões GomesAinda não há avaliações
- Sebenta Antiga AlunaDocumento196 páginasSebenta Antiga AlunaPlot BreakAinda não há avaliações
- MI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaDocumento4 páginasMI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaEriczinAinda não há avaliações
- Biologia e Geologia - 10º Ano - Obtenção de MatériaDocumento75 páginasBiologia e Geologia - 10º Ano - Obtenção de MatériaHugo MartinsAinda não há avaliações
- Membrana Celular: Características e FunçõesDocumento24 páginasMembrana Celular: Características e FunçõesAna Barth100% (1)
- Biologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDocumento25 páginasBiologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDouglas NobreAinda não há avaliações
- Membrana celular: transporte e permeabilidadeDocumento34 páginasMembrana celular: transporte e permeabilidadeGT Pericia FarmacêuticaAinda não há avaliações
- Membrana Plasmática: Transporte e EstruturasDocumento32 páginasMembrana Plasmática: Transporte e EstruturasPriscila MarinAinda não há avaliações
- Membrana Plasmática - BIOLOGIADocumento2 páginasMembrana Plasmática - BIOLOGIAJully ReisAinda não há avaliações
- Aula 1 - Biologia Celular e BioquímicaDocumento48 páginasAula 1 - Biologia Celular e BioquímicaFernando Paes Barreto AtrochAinda não há avaliações
- Aula 4 - Membrana PlasmáticaDocumento16 páginasAula 4 - Membrana PlasmáticaUrias NobreAinda não há avaliações
- A Estrutura e Funções da Membrana PlasmaticaDocumento7 páginasA Estrutura e Funções da Membrana PlasmaticaMT CRAFTAinda não há avaliações
- Membrana PlasmáticaDocumento6 páginasMembrana PlasmáticaLeticia KirchimairAinda não há avaliações
- Membrana+transportesDocumento49 páginasMembrana+transportesSara SilvaAinda não há avaliações
- Membrana Plasmática - SlidesDocumento41 páginasMembrana Plasmática - SlidesAnna BeatryzAinda não há avaliações
- Difusão e OsmoseDocumento46 páginasDifusão e OsmoseSerzhyo MashaduAinda não há avaliações
- Membrana Plasmática Transporte Funções PropriedadesDocumento37 páginasMembrana Plasmática Transporte Funções PropriedadesPalmeirizando TV100% (1)
- Biologia - Membrana Celular e Transportes TransmembranaresDocumento5 páginasBiologia - Membrana Celular e Transportes TransmembranaresJoão AzevedoAinda não há avaliações
- Biologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Documento65 páginasBiologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Leticia RodriguesAinda não há avaliações
- A estrutura e função da célulaDocumento8 páginasA estrutura e função da célulaKarina PezotiAinda não há avaliações
- Obtenção de matéria biológicaDocumento4 páginasObtenção de matéria biológicafrancisca soaresAinda não há avaliações
- UN3Documento13 páginasUN3Gildo Flório JúniorAinda não há avaliações
- Membrana PlasmáticaDocumento10 páginasMembrana PlasmáticaNz NaganAinda não há avaliações
- Biologia Celular pt1Documento22 páginasBiologia Celular pt1Rapha BuenoAinda não há avaliações
- Membrana celular permeabilidadeDocumento21 páginasMembrana celular permeabilidadeAriadne SiqueiraAinda não há avaliações
- Obtenção de nutrientes por seres heterotróficosDocumento21 páginasObtenção de nutrientes por seres heterotróficosgentedanaveAinda não há avaliações
- Membrana Plasmática e A PermeabilidadeDocumento39 páginasMembrana Plasmática e A PermeabilidadeWesleyJúniorAinda não há avaliações
- A estrutura e funções da membrana celularDocumento4 páginasA estrutura e funções da membrana celularcarolmarquesAinda não há avaliações
- Introdução Á Biologia Celular e Do DesenvolvimentoDocumento20 páginasIntrodução Á Biologia Celular e Do DesenvolvimentoThaynara TolentinoAinda não há avaliações
- Citologia MP e CDocumento10 páginasCitologia MP e CBird ToxicAinda não há avaliações
- Membrana plasmática: estrutura e funçõesDocumento4 páginasMembrana plasmática: estrutura e funçõesDuda TavaresAinda não há avaliações
- Os processos de obtenção de matéria em seres heterotróficosDocumento7 páginasOs processos de obtenção de matéria em seres heterotróficosSonjaAinda não há avaliações
- AULA 2 - Fisiologia Da MembranaDocumento25 páginasAULA 2 - Fisiologia Da MembranaPedro LucasAinda não há avaliações
- UFRA Membrana plasmática funções permeabilidadeDocumento46 páginasUFRA Membrana plasmática funções permeabilidadequel04Ainda não há avaliações
- Bases Da Biologia Celular e Envoltórios CelularesDocumento53 páginasBases Da Biologia Celular e Envoltórios Celularesaline.fernandes11Ainda não há avaliações
- Apostila de Biologia - Vol 2Documento38 páginasApostila de Biologia - Vol 2Biologia Concurso Vestibular100% (2)
- Obtenção de Matéria Por Seres HeterotróficosDocumento10 páginasObtenção de Matéria Por Seres HeterotróficosDD VitoldasAinda não há avaliações
- Membrana celular: estrutura e transporteDocumento7 páginasMembrana celular: estrutura e transportegentedanaveAinda não há avaliações
- Prova de Bio Cel ResumoDocumento8 páginasProva de Bio Cel ResumoThais GonzagaAinda não há avaliações
- Apostila - BioCelDocumento40 páginasApostila - BioCelcamilaAinda não há avaliações
- Aula Membrama P e Parede CDocumento33 páginasAula Membrama P e Parede CGeorge Da JosinaAinda não há avaliações
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDocumento7 páginasBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaAinda não há avaliações
- A03.U.I - Fisiologia Do Sistema TegumentarDocumento26 páginasA03.U.I - Fisiologia Do Sistema TegumentarSamuel AlvesAinda não há avaliações
- Enzimas EstéticaDocumento2 páginasEnzimas EstéticaIsabela FernandaAinda não há avaliações
- VRPO - Valores de procedimentos odontológicosDocumento2 páginasVRPO - Valores de procedimentos odontológicosKasuete Ono0% (2)
- Origem Vida Lista ExercíciosDocumento8 páginasOrigem Vida Lista ExercíciosbartolomeukumaAinda não há avaliações
- Crioterapia na reabilitação de lesõesDocumento52 páginasCrioterapia na reabilitação de lesõesMarcelo Barbosa da CruzAinda não há avaliações
- Sinais Vitais IbsDocumento11 páginasSinais Vitais IbsAmanda FerreiraAinda não há avaliações
- Encefalomalácia aviária 5o semestre UNILAGODocumento7 páginasEncefalomalácia aviária 5o semestre UNILAGORoberto RibeiroAinda não há avaliações
- Checklist Semiologia Cardíaca 2019 - 1Documento2 páginasChecklist Semiologia Cardíaca 2019 - 1Gloria DiasAinda não há avaliações
- Abordagem Do Paciênte Com DispnéiaDocumento4 páginasAbordagem Do Paciênte Com Dispnéiargp1987Ainda não há avaliações
- UFBA ICSB03 Teoria Mioelástica de Van Den BergDocumento8 páginasUFBA ICSB03 Teoria Mioelástica de Van Den BergMaria Amanda de Castro CâmaraAinda não há avaliações
- Guia prático para combater dores articulares sem anti-inflamatóriosDocumento18 páginasGuia prático para combater dores articulares sem anti-inflamatóriosJosé Marcos de OliveiraAinda não há avaliações
- Trabalho Paladar ProntoDocumento17 páginasTrabalho Paladar Prontonaycg50% (2)
- 46 Coleta de Exames LaboratoriasDocumento11 páginas46 Coleta de Exames LaboratoriasKelly Leite100% (1)
- Introdução à BotânicaDocumento178 páginasIntrodução à BotânicaPaula Cruz100% (1)
- Framboesa Morfologia e FisiologiaDocumento39 páginasFramboesa Morfologia e FisiologiaJoão CarraçaAinda não há avaliações
- Sistema EndócrinoDocumento18 páginasSistema EndócrinoAline ChristineAinda não há avaliações
- Atividade Avaliativa de Ciências Células e ÁguaDocumento1 páginaAtividade Avaliativa de Ciências Células e Águaciencia100% (1)
- Movimentos da cintura escapular e seus músculosDocumento23 páginasMovimentos da cintura escapular e seus músculosJRamilton Borges de BarrosAinda não há avaliações
- Psi Co Linguist Í CADocumento5 páginasPsi Co Linguist Í CASadraque CousinAinda não há avaliações
- Introdução à Anatomia HumanaDocumento13 páginasIntrodução à Anatomia HumanaBruno SoaresAinda não há avaliações
- Ebook Focus Project Ed - FinalDocumento37 páginasEbook Focus Project Ed - FinalBruno TeixeiraAinda não há avaliações
- Correção da Ficha de Exercícios de Biologia e GeologiaDocumento13 páginasCorreção da Ficha de Exercícios de Biologia e GeologiaFlávia FernandesAinda não há avaliações
- Mecanismo de Ação Dos FarmacosDocumento28 páginasMecanismo de Ação Dos FarmacosNatã DiasAinda não há avaliações
- Herdabilidade Genética em AnimaisDocumento32 páginasHerdabilidade Genética em AnimaisFabiane RibeiroAinda não há avaliações
- Tratamento de Rugas Dinâmicas Com EndermoterapiaDocumento41 páginasTratamento de Rugas Dinâmicas Com EndermoterapiaDebora Yoshie Iwata GarciaAinda não há avaliações
- Sistema respiratório e trocas gasosasDocumento4 páginasSistema respiratório e trocas gasosasPedro SousaAinda não há avaliações