Você também pode gostar
- Células eucariontes: estrutura e funçãoDocumento5 páginasCélulas eucariontes: estrutura e funçãoLucas NevesAinda não há avaliações
- Fechamento Problema 2Documento4 páginasFechamento Problema 2anna luisaAinda não há avaliações
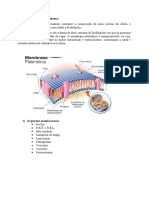
- Membrana CelularDocumento10 páginasMembrana CelularEsmeralda MuchisseAinda não há avaliações
- Estrutura das Membranas CelularesDocumento19 páginasEstrutura das Membranas CelularesThiago SantosAinda não há avaliações
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Documento13 páginasModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixAinda não há avaliações
- Compartimentos Intracelulares e TransporteDocumento8 páginasCompartimentos Intracelulares e Transportefevabax924Ainda não há avaliações
- Biologia - Membrana Celular e Transportes TransmembranaresDocumento5 páginasBiologia - Membrana Celular e Transportes TransmembranaresJoão AzevedoAinda não há avaliações
- Biologia Molecular Da Célula (Alberts Et Al, 5 Ed.)Documento13 páginasBiologia Molecular Da Célula (Alberts Et Al, 5 Ed.)Rodrigo SimaoAinda não há avaliações
- Resumo Biocel 1 Etapa - LuizaDocumento19 páginasResumo Biocel 1 Etapa - Luizaraf.marques.20Ainda não há avaliações
- Estrutura e função das membranas celulares e sistemas de transporteDocumento14 páginasEstrutura e função das membranas celulares e sistemas de transporteLuis RicardoAinda não há avaliações
- MI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaDocumento4 páginasMI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaEriczinAinda não há avaliações
- Apostila-Citologia-pg 19 A 34Documento16 páginasApostila-Citologia-pg 19 A 34denisetg100% (2)
- BCELDocumento12 páginasBCELMaria lAinda não há avaliações
- Membrana plasmática: estrutura e funçõesDocumento4 páginasMembrana plasmática: estrutura e funçõesDuda TavaresAinda não há avaliações
- Resumo ProliferaçãoDocumento41 páginasResumo ProliferaçãoRodrigo Augusto100% (8)
- HISTOLOGIADocumento18 páginasHISTOLOGIARebecca SicariAinda não há avaliações
- Aula MembranasDocumento33 páginasAula MembranasClaudio CirneAinda não há avaliações
- Resumo - GenéticaDocumento16 páginasResumo - Genéticaisabeljulia968Ainda não há avaliações
- Compartimentos Intracelulares e Endereçamento de ProteínasDocumento11 páginasCompartimentos Intracelulares e Endereçamento de ProteínasGabriel Da Silva SengesAinda não há avaliações
- Membrana PlasmáticaDocumento10 páginasMembrana PlasmáticaNz NaganAinda não há avaliações
- Membrana Plasmática: Transporte e EstruturasDocumento32 páginasMembrana Plasmática: Transporte e EstruturasPriscila MarinAinda não há avaliações
- Histologia - CitologiaDocumento12 páginasHistologia - CitologiaPhelipe MonteiroAinda não há avaliações
- Células: estrutura e função básicaDocumento33 páginasCélulas: estrutura e função básicaÂngela AlvesAinda não há avaliações
- Fisiologia VegetalDocumento8 páginasFisiologia VegetalRafael GuardadoAinda não há avaliações
- A estrutura e função da célulaDocumento8 páginasA estrutura e função da célulaKarina PezotiAinda não há avaliações
- Estudo Biologia2Documento9 páginasEstudo Biologia2WQWQWEQWEQAinda não há avaliações
- Membrana Celular: Transporte e Estrutura emDocumento46 páginasMembrana Celular: Transporte e Estrutura emTatiane ValeAinda não há avaliações
- Obtenção de Matéria PDFDocumento15 páginasObtenção de Matéria PDFLeticia RussoAinda não há avaliações
- Membrana celular: funções e estruturaDocumento20 páginasMembrana celular: funções e estruturaMario Maganizo JuniorAinda não há avaliações
- Matéria Orgânica PDFDocumento13 páginasMatéria Orgânica PDFLeticia RussoAinda não há avaliações
- Membrana Plasmática: Estrutura, Funções e TransporteDocumento5 páginasMembrana Plasmática: Estrutura, Funções e TransporteMoises FerroAinda não há avaliações
- Biologia Celular pt1Documento22 páginasBiologia Celular pt1Rapha BuenoAinda não há avaliações
- Resumo Complexo JuncionalDocumento5 páginasResumo Complexo JuncionalLuíza SiqueiraAinda não há avaliações
- Apostila de Biologia - Vol 2Documento38 páginasApostila de Biologia - Vol 2Biologia Concurso Vestibular100% (2)
- Biologia Transporte Na CélulaDocumento3 páginasBiologia Transporte Na CélulaMárcio Simões GomesAinda não há avaliações
- Citoesqueleto e CitoplasmaDocumento35 páginasCitoesqueleto e CitoplasmaRicardo GomesAinda não há avaliações
- Princípios de Neurociência - Kandel 5a EdDocumento40 páginasPrincípios de Neurociência - Kandel 5a EdcarlosAinda não há avaliações
- CitoesqueletoDocumento4 páginasCitoesqueletoCláudia MartinsAinda não há avaliações
- Biologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Documento65 páginasBiologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Leticia RodriguesAinda não há avaliações
- Membrana Plasmática - BIOLOGIADocumento2 páginasMembrana Plasmática - BIOLOGIAJully ReisAinda não há avaliações
- As células: unidades funcionais dos seres vivosDocumento10 páginasAs células: unidades funcionais dos seres vivosHálysson Nogueira ShalomAinda não há avaliações
- Psicobiologia PsiclinicaDocumento86 páginasPsicobiologia Psiclinicaneuropartilha4665Ainda não há avaliações
- Biologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDocumento25 páginasBiologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDouglas NobreAinda não há avaliações
- O papel dos filamentos do citoesqueletoDocumento8 páginasO papel dos filamentos do citoesqueletoAdamastor NetoAinda não há avaliações
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDocumento7 páginasBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaAinda não há avaliações
- Estudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasDocumento2 páginasEstudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasLuiza LopesAinda não há avaliações
- Fichas - Organelas CitoplasmáticasDocumento6 páginasFichas - Organelas CitoplasmáticasEstefania RoblesAinda não há avaliações
- Especializações Da Membrana PlasmáticaDocumento18 páginasEspecializações Da Membrana PlasmáticaPaula AlmeidaAinda não há avaliações
- Resumos de Biologia CelularDocumento17 páginasResumos de Biologia CelularCaroline LimaAinda não há avaliações
- Componentes celularesDocumento17 páginasComponentes celularesVera DominguesAinda não há avaliações
- Fisiologia celular: estrutura e função das célulasDocumento15 páginasFisiologia celular: estrutura e função das célulasAlícia Hellen100% (1)
- Membrana Celular: Características e FunçõesDocumento24 páginasMembrana Celular: Características e FunçõesAna Barth100% (1)
- A estrutura e função da membrana plasmáticaDocumento2 páginasA estrutura e função da membrana plasmáticaMaria Theresa MoreiraAinda não há avaliações
- Membrana Plasmática. Digestão IntracelularDocumento27 páginasMembrana Plasmática. Digestão IntracelularBruno PereiraAinda não há avaliações
- BiocelDocumento23 páginasBiocelSalvar Meus JoguinhosAinda não há avaliações
- Doenças Biologia1Documento2 páginasDoenças Biologia1ti18Ainda não há avaliações
- Gripes No MundoDocumento1 páginaGripes No MundoThais GonzagaAinda não há avaliações
- Conselhos Úteis para Quem Vai Fazer A Cidadania Italiana Na Itália - Antes de Viajar Cidadania ItalianaDocumento3 páginasConselhos Úteis para Quem Vai Fazer A Cidadania Italiana Na Itália - Antes de Viajar Cidadania ItalianaThais GonzagaAinda não há avaliações
- Trabalho Da Exposição Mario TestinoDocumento1 páginaTrabalho Da Exposição Mario TestinoThais GonzagaAinda não há avaliações
- Cheesecake CopinhoDocumento1 páginaCheesecake CopinhoThais GonzagaAinda não há avaliações
- Atraente Alma-Brasileira FlautaDocumento4 páginasAtraente Alma-Brasileira FlautaThais GonzagaAinda não há avaliações
- Mousse de Chocolate fácilDocumento1 páginaMousse de Chocolate fácilThais GonzagaAinda não há avaliações
- CandvagaDocumento7 páginasCandvagaAndreza Silveira PereiraAinda não há avaliações
- Exercicios Propostos Condensadores-Evaporadores Prof. Van RaphDocumento32 páginasExercicios Propostos Condensadores-Evaporadores Prof. Van RaphVAN GOMPEL RAPH100% (1)
- Otro Tipo de SupositoriosDocumento23 páginasOtro Tipo de SupositoriosYoxe MolinaAinda não há avaliações
- 3 ExercResolv e Exerc 3.4 PooleDocumento5 páginas3 ExercResolv e Exerc 3.4 PoolelataodeprasticoAinda não há avaliações
- Notas de Aula de HidráulicaDocumento138 páginasNotas de Aula de HidráulicaJéssica OliveiraAinda não há avaliações
- O que é Sistema GeodésicoDocumento12 páginasO que é Sistema GeodésicoSávio Gouvêa de Freitas100% (1)
- Controle térmico aumenta rendimento de carvãoDocumento12 páginasControle térmico aumenta rendimento de carvãoCrismeire IsbaexAinda não há avaliações
- @ligaliteraria Caleidoscopio - Sue Hecker PDFDocumento417 páginas@ligaliteraria Caleidoscopio - Sue Hecker PDFmoises azevedoAinda não há avaliações
- Laudo de Verificação de Patologias Construtivas Causadas Por Obra LindeiraDocumento21 páginasLaudo de Verificação de Patologias Construtivas Causadas Por Obra LindeiraClarel Da Cruz RietAinda não há avaliações
- Motores Diesel 2.5l TDI da Volkswagen CrafterDocumento38 páginasMotores Diesel 2.5l TDI da Volkswagen CrafterIgatech Diesel100% (2)
- Aula 5 - Instrumentos HidrometeorológicosDocumento69 páginasAula 5 - Instrumentos Hidrometeorológicospedroi17100% (1)
- MANUAL DA GELADEIRA DF38 - Eletrolux PDFDocumento36 páginasMANUAL DA GELADEIRA DF38 - Eletrolux PDFCristina Tomaschewsky67% (6)
- Como usar PhotoFiltreDocumento12 páginasComo usar PhotoFiltrejujacarneiroAinda não há avaliações
- Manual Gerador Gasolina GG 12000 - V1Documento28 páginasManual Gerador Gasolina GG 12000 - V1José Nilton Diniz FranqueiraAinda não há avaliações
- Motores InduçãoDocumento3 páginasMotores InduçãoMariana AndradeAinda não há avaliações
- Construção Com Solo CimentoDocumento3 páginasConstrução Com Solo CimentoSandersonAinda não há avaliações
- CabosDocumento26 páginasCabosreinscAinda não há avaliações
- Análise estatística da Mega SenaDocumento22 páginasAnálise estatística da Mega Senaprophecta100% (3)
- Bomba HidrárulicasDocumento56 páginasBomba HidrárulicasJocelia LopesAinda não há avaliações
- O PSICO GERAD-WPS OfficeDocumento8 páginasO PSICO GERAD-WPS OfficeLuis Carlos Almeida100% (2)
- Método de CrossDocumento14 páginasMétodo de CrossGADDC100% (1)
- Tese Completa Oliveira (2004)Documento111 páginasTese Completa Oliveira (2004)ClarissaAinda não há avaliações
- Manual Uso Sistema KNXDocumento27 páginasManual Uso Sistema KNXHélderAinda não há avaliações
- Materiais - ParafusosDocumento56 páginasMateriais - ParafusosPaulo S. T. Vanucci0% (1)
- Prova 4 - Moço de ConvésDocumento9 páginasProva 4 - Moço de ConvésJose Carlos FernandesAinda não há avaliações
- LT - Aula01 - Introdução A Linhas de TransmissãoDocumento30 páginasLT - Aula01 - Introdução A Linhas de TransmissãoDionatan RangelAinda não há avaliações
- MANUAL BOBINAGEM MOTOR WEG TRIFÁSICODocumento3 páginasMANUAL BOBINAGEM MOTOR WEG TRIFÁSICOSeverino OviedoAinda não há avaliações
- Vectores e suas operaçõesDocumento26 páginasVectores e suas operaçõesSandra Cristina da Silva FernandesAinda não há avaliações
- A Pequena Sereia e Os Reinos Das Ilusoes PDFDocumento258 páginasA Pequena Sereia e Os Reinos Das Ilusoes PDFMay RS7Ainda não há avaliações
- Ferramentas de Corte UsinagemDocumento4 páginasFerramentas de Corte UsinagemFrajolaAinda não há avaliações
- Os NúmerosDocumento171 páginasOs NúmerosAPPARATOR100% (29)