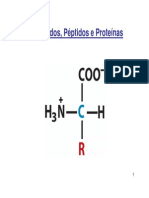
Você também pode gostar
- Membrana PlasmáticaDocumento44 páginasMembrana PlasmáticaThaynara TolentinoAinda não há avaliações
- Estrutura e funções da membrana celularDocumento10 páginasEstrutura e funções da membrana celularJulia AraujoAinda não há avaliações
- Membranas CelularesDocumento22 páginasMembranas Celularesluisa diasAinda não há avaliações
- Membrana celular: funções e estruturaDocumento20 páginasMembrana celular: funções e estruturaMario Maganizo JuniorAinda não há avaliações
- PlementarDocumento3 páginasPlementarelizangelacarAinda não há avaliações
- Membrana Plasmatica BioquimDocumento83 páginasMembrana Plasmatica Bioquimflavia douraoAinda não há avaliações
- Resumo Biocel 1 Etapa - LuizaDocumento19 páginasResumo Biocel 1 Etapa - Luizaraf.marques.20Ainda não há avaliações
- Envoltorios CelularesDocumento41 páginasEnvoltorios CelularesJonhy CleberAinda não há avaliações
- Aula 2 Biofísica 24Documento22 páginasAula 2 Biofísica 24Sílvia ScabinAinda não há avaliações
- Estudo Dirigido 2 - CitologiaDocumento4 páginasEstudo Dirigido 2 - CitologiaYara KerberAinda não há avaliações
- Estrutura e composição das membranas biológicasDocumento5 páginasEstrutura e composição das membranas biológicasLuara FontesAinda não há avaliações
- BIOMEMBRANASDocumento4 páginasBIOMEMBRANASaninhayomeAinda não há avaliações
- Membrana Celular: Transporte e Modelo Mosaico-FluidoDocumento8 páginasMembrana Celular: Transporte e Modelo Mosaico-FluidoMatilde PedroAinda não há avaliações
- Aula 4 - Resumo - 2009Documento6 páginasAula 4 - Resumo - 2009Isabela LiaAinda não há avaliações
- Membrana celular permeabilidadeDocumento21 páginasMembrana celular permeabilidadeAriadne SiqueiraAinda não há avaliações
- Membrana Celular: Características e FunçõesDocumento24 páginasMembrana Celular: Características e FunçõesAna Barth100% (1)
- Apostila - BioCelDocumento40 páginasApostila - BioCelcamilaAinda não há avaliações
- BiocelDocumento23 páginasBiocelSalvar Meus JoguinhosAinda não há avaliações
- Membranas CelularesDocumento8 páginasMembranas CelularesMarcela BrasilAinda não há avaliações
- Membrana PlasmáticaDocumento42 páginasMembrana PlasmáticaMelina MarquesAinda não há avaliações
- A Estrutura e Funções da Membrana PlasmaticaDocumento7 páginasA Estrutura e Funções da Membrana PlasmaticaMT CRAFTAinda não há avaliações
- Aula 1. BiomembranasDocumento33 páginasAula 1. BiomembranasFUTSHOW CATANDUVAAinda não há avaliações
- Biologia e Geologia - 10º Ano - Obtenção de MatériaDocumento75 páginasBiologia e Geologia - 10º Ano - Obtenção de MatériaHugo MartinsAinda não há avaliações
- Histologia - CitologiaDocumento12 páginasHistologia - CitologiaPhelipe MonteiroAinda não há avaliações
- Fechamento Problema 2Documento4 páginasFechamento Problema 2anna luisaAinda não há avaliações
- Membrana Plasmática: Transporte e EstruturasDocumento32 páginasMembrana Plasmática: Transporte e EstruturasPriscila MarinAinda não há avaliações
- BiomembranasDocumento15 páginasBiomembranasmati cAinda não há avaliações
- Membrana Biológica Mosaico FluidoDocumento87 páginasMembrana Biológica Mosaico FluidoVitória VignoliAinda não há avaliações
- Material de Apoio - Membrana Plasmática-Movimentação de Substâncias No Ambiente CelularDocumento14 páginasMaterial de Apoio - Membrana Plasmática-Movimentação de Substâncias No Ambiente CelularcitrolidaniAinda não há avaliações
- Biocel - PHDocumento70 páginasBiocel - PHpaulohnunesAinda não há avaliações
- Membrana Plasmatica 1 AnosDocumento16 páginasMembrana Plasmatica 1 AnosAna SilvaAinda não há avaliações
- Apostila-Citologia-pg 19 A 34Documento16 páginasApostila-Citologia-pg 19 A 34denisetg100% (2)
- Mon Bio Cel Ed Membrana RespDocumento5 páginasMon Bio Cel Ed Membrana RespRyan VieiraAinda não há avaliações
- Características e funções das membranas biológicasDocumento12 páginasCaracterísticas e funções das membranas biológicasAna Clara FrotaAinda não há avaliações
- Membrana PlasmáticaDocumento8 páginasMembrana PlasmáticaAna Julia PessotiAinda não há avaliações
- AULA 4 BIOLOGIA CELULAR Estrutura Celular Membrana Plasmática Citoplasma NúcleoDocumento41 páginasAULA 4 BIOLOGIA CELULAR Estrutura Celular Membrana Plasmática Citoplasma Núcleocaroldsll14Ainda não há avaliações
- Membranas 2016.1Documento50 páginasMembranas 2016.1Deusiane MuruciAinda não há avaliações
- Estrutura das Membranas CelularesDocumento19 páginasEstrutura das Membranas CelularesThiago SantosAinda não há avaliações
- Pink Illustrative Organic Biology Project PresentationDocumento21 páginasPink Illustrative Organic Biology Project PresentationJoão VitorAinda não há avaliações
- Estrutura e Funções da Membrana CelularDocumento3 páginasEstrutura e Funções da Membrana CelularMariana Gomes da Silva100% (1)
- Bases 1Documento34 páginasBases 1Thayná CrisóstomoAinda não há avaliações
- Captura de Tela 2023-04-10 À(s) 05.38.45Documento54 páginasCaptura de Tela 2023-04-10 À(s) 05.38.45Nicole Ruiz RolimAinda não há avaliações
- Estrutura Da MembranaDocumento5 páginasEstrutura Da MembranaVitor GabrielAinda não há avaliações
- 2 - Membrana CelularDocumento10 páginas2 - Membrana CelularJoana SangoAinda não há avaliações
- Biologia Celular IntroduçãoDocumento38 páginasBiologia Celular IntroduçãoMaxwell M. RibeiroAinda não há avaliações
- Membrana plasmática e transporteDocumento59 páginasMembrana plasmática e transporteRafael RezendeAinda não há avaliações
- Membrana CelularDocumento3 páginasMembrana CelularRodrigo KerberAinda não há avaliações
- AULA 3 BIOLOGIA CELULAR - Membranas Biologicas-1Documento81 páginasAULA 3 BIOLOGIA CELULAR - Membranas Biologicas-1Levi CamoesAinda não há avaliações
- Membrana celular e permeabilidadeDocumento33 páginasMembrana celular e permeabilidadehendrya08Ainda não há avaliações
- Conceitos Gerais das Membranas CelularesDocumento8 páginasConceitos Gerais das Membranas CelularesDaniela GondimAinda não há avaliações
- Membrana Plasmática: Estrutura, Funções e TransporteDocumento5 páginasMembrana Plasmática: Estrutura, Funções e TransporteMoises FerroAinda não há avaliações
- Estrutura da membrana21Documento6 páginasEstrutura da membrana21Monica AlexandreAinda não há avaliações
- Membrana Plasmática - SlidesDocumento41 páginasMembrana Plasmática - SlidesAnna BeatryzAinda não há avaliações
- Revisao para Prova de CitologiaDocumento30 páginasRevisao para Prova de CitologiaAdilson Marcos de SouzaAinda não há avaliações
- Membrana plasmática - biocelDocumento4 páginasMembrana plasmática - biocellarissazimmermann8Ainda não há avaliações
- CitologiaDocumento8 páginasCitologiaericc-13hotmail.comAinda não há avaliações
- BCM-unidade III 1-2Documento86 páginasBCM-unidade III 1-2Edmond TAinda não há avaliações
- CONCEITOS ESSENCIAIS Por Alberts Et AlDocumento2 páginasCONCEITOS ESSENCIAIS Por Alberts Et AlElias GallinaAinda não há avaliações
- Revista de BiologiaDocumento592 páginasRevista de BiologiaRamalde Santana100% (2)
- Células eucariontes: características e organelasDocumento9 páginasCélulas eucariontes: características e organelasluis augusto aquatti capatoAinda não há avaliações
- Dicionário de Genética e GATTACA - Experiência GenéticaDocumento3 páginasDicionário de Genética e GATTACA - Experiência GenéticaRodrigoAinda não há avaliações
- A Composição Química Da CélulaDocumento18 páginasA Composição Química Da CélulaMiguel RamosAinda não há avaliações
- Telómeros e envelhecimento celularDocumento8 páginasTelómeros e envelhecimento celularCatarina SáAinda não há avaliações
- Provas BiologiaDocumento22 páginasProvas BiologiaTeresa silva0% (1)
- Enzimas: Uma breve introdução às enzimasDocumento19 páginasEnzimas: Uma breve introdução às enzimasjackspolAinda não há avaliações
- Funções das organelas citoplasmáticasDocumento2 páginasFunções das organelas citoplasmáticasDarlan HenriqueAinda não há avaliações
- Componentes orgânicos e inorgânicos das célulasDocumento60 páginasComponentes orgânicos e inorgânicos das célulasjoice vianaAinda não há avaliações
- Mapa MetabolicoDocumento2 páginasMapa MetabolicoLuana A. BolzaniAinda não há avaliações
- Exercicios Bioquimica DPDocumento8 páginasExercicios Bioquimica DPjuliliossi100% (1)
- 2aula Aminoacidos (Modo de Compatibilidade)Documento38 páginas2aula Aminoacidos (Modo de Compatibilidade)Elizael De Jesus GonçalvesAinda não há avaliações
- Biologia molecular: dogma central e técnicasDocumento2 páginasBiologia molecular: dogma central e técnicasJanderson SalustianoAinda não há avaliações
- Fitatos e OxalatosDocumento2 páginasFitatos e OxalatosIsabel SerralheiroAinda não há avaliações
- Novo Intradermoterapia Resultados 2019Documento422 páginasNovo Intradermoterapia Resultados 2019Reijane SilvaAinda não há avaliações
- Biópsia de vilo corial e amniocentese: intervenções no pré-natalDocumento17 páginasBiópsia de vilo corial e amniocentese: intervenções no pré-natalMAISAMSBAinda não há avaliações
- Engenharia genética técnicas DNADocumento18 páginasEngenharia genética técnicas DNAAndresa SampaioAinda não há avaliações
- Proteínas: estrutura, propriedades e migração em campo elétricoDocumento3 páginasProteínas: estrutura, propriedades e migração em campo elétricoLuan SantosAinda não há avaliações
- Bionergética e Substratos EnergéticosDocumento57 páginasBionergética e Substratos EnergéticosVivian Tezzelle83% (6)
- Biologia 10ºanoDocumento15 páginasBiologia 10ºanoAdriana CandeiasAinda não há avaliações
- Ebook - Descomplicando A Bioquimica PDFDocumento101 páginasEbook - Descomplicando A Bioquimica PDFEdmara Silva100% (2)
- Cib 603 BiocelularDocumento2 páginasCib 603 BiocelularTalesAinda não há avaliações
- Lamina - MeiviewDocumento2 páginasLamina - MeiviewPaulaGiselleCzaikoskiAinda não há avaliações
- Roteiro de Aula Prática - "Cebola"Documento3 páginasRoteiro de Aula Prática - "Cebola"Gabriel teixeiraAinda não há avaliações
- Genômica Funcional e Epidemiologia GenéticaDocumento14 páginasGenômica Funcional e Epidemiologia Genéticarack26Ainda não há avaliações
- APANHADO COMPLETO BIOQUÍMICA UNIP 2023 - Passei Direto Parte 3Documento10 páginasAPANHADO COMPLETO BIOQUÍMICA UNIP 2023 - Passei Direto Parte 3avataresdacuraAinda não há avaliações
- Bioquímica Aplicada IIDocumento75 páginasBioquímica Aplicada IIjackspolAinda não há avaliações
- @vetezoo - BIOQUÍMICA CLÍNICA VETERINÁRIA 1Documento36 páginas@vetezoo - BIOQUÍMICA CLÍNICA VETERINÁRIA 1Maria Raquel SilvaAinda não há avaliações
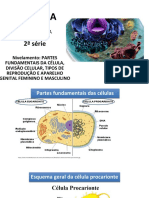
- Partes fundamentais da célula, divisão celular e tipos de reproduçãoDocumento22 páginasPartes fundamentais da célula, divisão celular e tipos de reproduçãoJoão Vitor S MachadoAinda não há avaliações
- Espécies reativas de oxigênio e sistemas antioxidantesDocumento5 páginasEspécies reativas de oxigênio e sistemas antioxidantesIsadora ScopelAinda não há avaliações
- Simetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaNo EverandSimetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaAinda não há avaliações
- Série Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNo EverandSérie Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNota: 5 de 5 estrelas5/5 (1)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Equilíbrio iônico: aplicações em química analíticaNo EverandEquilíbrio iônico: aplicações em química analíticaAinda não há avaliações
- Para Gostar de Química: Introdução à Química GeralNo EverandPara Gostar de Química: Introdução à Química GeralAinda não há avaliações
- Problemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)No EverandProblemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)Ainda não há avaliações
- Química da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaNo EverandQuímica da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaAinda não há avaliações
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Detetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseNo EverandDetetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseAinda não há avaliações
- Uso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaNo EverandUso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaAinda não há avaliações
- Vigilância Sanitária Nos Estados E MunicípiosNo EverandVigilância Sanitária Nos Estados E MunicípiosAinda não há avaliações
- Ácidos e Bases de Brönsted e Lowry: Uma visão aplicada ao meio ambienteNo EverandÁcidos e Bases de Brönsted e Lowry: Uma visão aplicada ao meio ambienteAinda não há avaliações
- Princípios de química analítica: Abordagem teórica qualitativa e quantitativaNo EverandPrincípios de química analítica: Abordagem teórica qualitativa e quantitativaNota: 5 de 5 estrelas5/5 (1)
- Química na Educação Básica: Ferramentas teóricas e práticasNo EverandQuímica na Educação Básica: Ferramentas teóricas e práticasNota: 4 de 5 estrelas4/5 (1)
- Físico-química de Blendas e Compósitos PoliméricosNo EverandFísico-química de Blendas e Compósitos PoliméricosNota: 1 de 5 estrelas1/5 (1)