Você também pode gostar
- 55bfebd7 33 2 Helixor Pratica Clinica PDFDocumento8 páginas55bfebd7 33 2 Helixor Pratica Clinica PDFUana MattosAinda não há avaliações
- Questões de Neuroanatomia - GabaritoDocumento5 páginasQuestões de Neuroanatomia - GabaritoArthur FreitasAinda não há avaliações
- Caso EstamiraDocumento4 páginasCaso EstamiraMavi PsiAinda não há avaliações
- Panorama das doenças negligenciadas: um estudo de caso in vitro e in vivo de compostos Químicos Naturais frente ao Tripanossoma cruziNo EverandPanorama das doenças negligenciadas: um estudo de caso in vitro e in vivo de compostos Químicos Naturais frente ao Tripanossoma cruziAinda não há avaliações
- Giorgi e BardinDocumento10 páginasGiorgi e BardinMarcelo Vial RoeheAinda não há avaliações
- Vias NeuraisDocumento13 páginasVias NeuraisGabriella DiasAinda não há avaliações
- Captura de Tela 2022-11-10 À(s) 01.13.49Documento19 páginasCaptura de Tela 2022-11-10 À(s) 01.13.49Rayane LopesAinda não há avaliações
- CerebeloDocumento10 páginasCerebeloclaraAinda não há avaliações
- Material Apoio Sistema Nervoso GABARITODocumento3 páginasMaterial Apoio Sistema Nervoso GABARITOLuiza Brandino100% (2)
- 09289P - Análise Textual DiscursivaDocumento2 páginas09289P - Análise Textual DiscursivaWiliam RomanholiAinda não há avaliações
- Sistema LimbicoDocumento46 páginasSistema LimbicoGabriel RibeiroAinda não há avaliações
- Avaliação FinalDocumento6 páginasAvaliação FinalGuiomar Santos Da SilvaAinda não há avaliações
- Insônia No IdosoDocumento16 páginasInsônia No IdosoFábio Vinholy100% (1)
- Resumo de Neurofisiologia (Psicologia)Documento7 páginasResumo de Neurofisiologia (Psicologia)Jeniffer Fernandes100% (1)
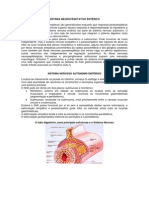
- Sistema Nervoso AutônomoDocumento12 páginasSistema Nervoso AutônomoIzadora Dias Bessa de AlmeidaAinda não há avaliações
- Estudo Dirigido 3 - AmebasDocumento3 páginasEstudo Dirigido 3 - AmebasNicolly BertassoAinda não há avaliações
- Febre Amarela No Brasil FGV 39 PG PDFDocumento39 páginasFebre Amarela No Brasil FGV 39 PG PDFAdalberto BatistaAinda não há avaliações
- Cerebelo (Resumo)Documento4 páginasCerebelo (Resumo)Fábio Constancio Almeida da Silva100% (1)
- Genética Dos Grupos Sanguíneos FARMDocumento11 páginasGenética Dos Grupos Sanguíneos FARMGabriel Isaac BelliniAinda não há avaliações
- Neurônios Monoaminérgicos Do Tronco EncefálicoDocumento4 páginasNeurônios Monoaminérgicos Do Tronco EncefálicoLucas Freitas100% (1)
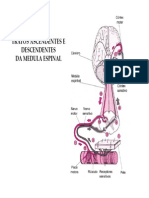
- Tratos Ascendentes e Descendentes Da ME (Modo de Compatibilidade)Documento13 páginasTratos Ascendentes e Descendentes Da ME (Modo de Compatibilidade)Natália PimentelAinda não há avaliações
- Estudo Dirigido Histórico - JenniferDocumento3 páginasEstudo Dirigido Histórico - JenniferJennifer GuimaraesAinda não há avaliações
- 5 SomestesiaDocumento34 páginas5 SomestesiaElisangela Aquino100% (1)
- No caminho certo após uma cirurgia cardíaca: Um guia completo com orientações e recomendações para o pós-operatórioNo EverandNo caminho certo após uma cirurgia cardíaca: Um guia completo com orientações e recomendações para o pós-operatórioAinda não há avaliações
- Wilhelm WundtDocumento7 páginasWilhelm WundtCarlos Adriano SilvaAinda não há avaliações
- Desvendando Os Segredos Dos Laboratórios: A Importância Das Análises Clínicas Na Saúde E Diagnóstico De DoençasNo EverandDesvendando Os Segredos Dos Laboratórios: A Importância Das Análises Clínicas Na Saúde E Diagnóstico De DoençasAinda não há avaliações
- Resumo Neurofisiologia Clinica Principios Basicos e Aplicacoes Luiz Carlos PintoDocumento2 páginasResumo Neurofisiologia Clinica Principios Basicos e Aplicacoes Luiz Carlos PintoJosehp Millares TolaAinda não há avaliações
- Princípios Da Neurotransmissão Química: Dra. Meiriélle Martins Meneghini PsiquiatriaDocumento55 páginasPrincípios Da Neurotransmissão Química: Dra. Meiriélle Martins Meneghini PsiquiatriaDeyvid Dos AnjosAinda não há avaliações
- AULA 2 Neronio Glia SinapseDocumento34 páginasAULA 2 Neronio Glia SinapseCamila Amorim100% (2)
- Neuroanatomia FuncionalDocumento10 páginasNeuroanatomia FuncionalDébora SouzaAinda não há avaliações
- REsumo Cap 4 e 5 Do Livro A Entrevista de AjudaDocumento1 páginaREsumo Cap 4 e 5 Do Livro A Entrevista de AjudaMarcelo MarchioriAinda não há avaliações
- 06 - Sistema LímbicoDocumento14 páginas06 - Sistema LímbicoDrksBrAinda não há avaliações
- Neuroanatomofisiologia - Perguntas UnificadasDocumento4 páginasNeuroanatomofisiologia - Perguntas UnificadasEduardo PK MartinsAinda não há avaliações
- 5f6ba90448932 - Macroscopia Do TelencefaloDocumento23 páginas5f6ba90448932 - Macroscopia Do TelencefaloGabriel FAinda não há avaliações
- Lucas Spanemberg Aula 3Documento119 páginasLucas Spanemberg Aula 3anathaiseAinda não há avaliações
- Sistema NervosoDocumento41 páginasSistema NervosoJoão LucasAinda não há avaliações
- Medicina LegalDocumento8 páginasMedicina Legaldouglas ramosAinda não há avaliações
- Rev PsiquiatriaDocumento56 páginasRev PsiquiatriacayonamineAinda não há avaliações
- 15 - Estrutura e Funções Dos Núcleos Da BaseDocumento10 páginas15 - Estrutura e Funções Dos Núcleos Da BaseReinaldo RiquetAinda não há avaliações
- Anatomia e Fisiologia Do Sistema NervosoDocumento31 páginasAnatomia e Fisiologia Do Sistema NervosoHenrique Araujo100% (1)
- Sistema Esquelético 21.06.2021Documento47 páginasSistema Esquelético 21.06.2021Kateriny BispoAinda não há avaliações
- Puberdade PrecoceDocumento7 páginasPuberdade PrecocetathiAinda não há avaliações
- 2022 - Aula Sistema NervosoDocumento33 páginas2022 - Aula Sistema NervosoEduardo Ananias100% (1)
- Ensaio Acerca Dos Sintomas SimbolicoDocumento129 páginasEnsaio Acerca Dos Sintomas SimbolicomairrconAinda não há avaliações
- Neuroanatomia - Completa (Atualizada 08-02-2012) High ResolutionDocumento184 páginasNeuroanatomia - Completa (Atualizada 08-02-2012) High ResolutionAlessandra Zapelloni100% (3)
- Cerebelo - Morfologia e FunçõesDocumento2 páginasCerebelo - Morfologia e FunçõesJediel PedrosoAinda não há avaliações
- Citologia ActinomycesDocumento2 páginasCitologia ActinomycesRaquel CostaAinda não há avaliações
- Miastenia GravisDocumento18 páginasMiastenia Gravisanasuhet100% (2)
- 08 Resumo - Lobos Cerebrais, Estruturas e Funçoes PDFDocumento3 páginas08 Resumo - Lobos Cerebrais, Estruturas e Funçoes PDFVinicius HenriqueAinda não há avaliações
- Sindrome Igg4Documento72 páginasSindrome Igg4Hugo Nogueira100% (1)
- Perfil e Prevalência Dos Tumores Primários Do Sistema NervosoDocumento6 páginasPerfil e Prevalência Dos Tumores Primários Do Sistema NervosoAllan MaiaAinda não há avaliações
- Capovilla e Garcia, LInguagem e CogniçãoDocumento320 páginasCapovilla e Garcia, LInguagem e CogniçãoVeronica GurgelAinda não há avaliações
- 1-Evolucao BiologicaDocumento83 páginas1-Evolucao BiologicaRafael AraujoAinda não há avaliações
- Características Da Citologia BacterianaDocumento3 páginasCaracterísticas Da Citologia BacterianaFabíola Araújo Barboza CoimbraAinda não há avaliações
- Resumo - Sistema Nervoso PeriféricoDocumento2 páginasResumo - Sistema Nervoso PeriféricoAmanda FrancoAinda não há avaliações
- MicrocirculaçãoDocumento5 páginasMicrocirculaçãoGuilherme Secundini de Almeida100% (1)
- Sistema Nervoso MiotentericoDocumento20 páginasSistema Nervoso MiotentericoJorge CamargoAinda não há avaliações
- Sistema Neurovegetativo EntéricoDocumento4 páginasSistema Neurovegetativo EntéricoSheila K. R. MargrafAinda não há avaliações
- Anatomia Unidade 3Documento20 páginasAnatomia Unidade 3antoniel100Ainda não há avaliações
- NutrologiaDocumento2 páginasNutrologiaThiago B.Ainda não há avaliações
- Edital 2021 08 Agente Censitario de Administracao e InformaticaDocumento132 páginasEdital 2021 08 Agente Censitario de Administracao e InformaticaMaria SilvaAinda não há avaliações
- 5 Gestao Da Qualidade em Servicos de SaudeDocumento9 páginas5 Gestao Da Qualidade em Servicos de SaudeThatiana GalvãoAinda não há avaliações
- Angio CranioDocumento4 páginasAngio CranioWesley CastroAinda não há avaliações
- Nutrição & MitosDocumento37 páginasNutrição & MitosDiogo JuniorAinda não há avaliações
- TERMO ADITIVO ESTÁGIO OrientaçõesDocumento1 páginaTERMO ADITIVO ESTÁGIO OrientaçõesMauricio BarrosAinda não há avaliações
- Relatorio DisplasiaDocumento5 páginasRelatorio DisplasiaMarianasouza SouzaAinda não há avaliações
- PSICOPATOLOGIA 3correçãoDocumento3 páginasPSICOPATOLOGIA 3correçãoAriane GomesAinda não há avaliações
- Ficha de Inscrição - Cfaq Maf-Mma n1 - Extra FdpemDocumento1 páginaFicha de Inscrição - Cfaq Maf-Mma n1 - Extra FdpemLuiz de Oliveira GomesAinda não há avaliações
- 03 - MAR - 09, 10 e 11 - SEC PMARDocumento6 páginas03 - MAR - 09, 10 e 11 - SEC PMARLuiza GamaAinda não há avaliações
- CLArtt Ca Ce Sup Sent 2019Documento17 páginasCLArtt Ca Ce Sup Sent 2019Daniele SáAinda não há avaliações
- Portfólio Fratura Óssea Parte 2 - Thamires LopesDocumento23 páginasPortfólio Fratura Óssea Parte 2 - Thamires LopesThamires Lopes100% (1)
- Sessão 7Documento48 páginasSessão 7Alves Neves SacaquiquiAinda não há avaliações
- Agua Dos MilagresDocumento3 páginasAgua Dos MilagreslkarolinoAinda não há avaliações
- 12 PressoterapiaDocumento38 páginas12 PressoterapiaJosiane RodriguesAinda não há avaliações
- 10sobremordida ExageradaDocumento12 páginas10sobremordida ExageradaLuana GuaitoliniAinda não há avaliações
- Método de Bligh-DyerDocumento3 páginasMétodo de Bligh-DyerFelipe ChicrallaAinda não há avaliações
- Caso ClinicoDocumento31 páginasCaso ClinicoFernandaCortezDonosoAinda não há avaliações
- Cap. 15 - CoccídiosDocumento17 páginasCap. 15 - CoccídiosPaulo CarvalhoAinda não há avaliações
- APR - PSO - Inspeção e Adequação de Painéis Elétricos e DisjuntoresDocumento3 páginasAPR - PSO - Inspeção e Adequação de Painéis Elétricos e DisjuntoresEzequiel Sorrentino de AzevedoAinda não há avaliações
- Saúde Da CriançaDocumento25 páginasSaúde Da CriançaMaurício FernandesAinda não há avaliações
- (Ferreira, Et Al., 2005) - Jurnal TinnitusDocumento8 páginas(Ferreira, Et Al., 2005) - Jurnal TinnitusHery PurnawantAinda não há avaliações
- Autismo Cartilha TeaDocumento15 páginasAutismo Cartilha TeaCPSST100% (5)
- Gestantes e SusDocumento13 páginasGestantes e SusPsiquiatria TranslacionalAinda não há avaliações
- Atlass de Parasitologia-TrematodosDocumento38 páginasAtlass de Parasitologia-TrematodosQUIMICO CLINICO WILLIANS SANCHEZ100% (2)
- Aula 1Documento37 páginasAula 1Jamylle GuedesAinda não há avaliações
- RPG InformativoDocumento1 páginaRPG InformativoGrazielle OliveiraAinda não há avaliações
- Clínica PET-FRIENDLY TPDocumento7 páginasClínica PET-FRIENDLY TPMadalena PimentaAinda não há avaliações
- Apostila - Sistema de Gestão - Faculdade Pitagoras PDFDocumento17 páginasApostila - Sistema de Gestão - Faculdade Pitagoras PDFmiaAinda não há avaliações
- Pos-Tratamento de Efluentes de Reatores AnaerobiosDocumento406 páginasPos-Tratamento de Efluentes de Reatores Anaerobiosjcbobeda100% (1)