Você também pode gostar
- Caso Clínico BlindagemDocumento11 páginasCaso Clínico BlindagemArmando JúniorAinda não há avaliações
- Caso Clínico BlindagemDocumento11 páginasCaso Clínico BlindagemArmando JúniorAinda não há avaliações
- Guia de endodontia laboratorialDocumento31 páginasGuia de endodontia laboratorialArmando JúniorAinda não há avaliações
- Versao Gratis - Apostila SUS para Concursos 2017Documento73 páginasVersao Gratis - Apostila SUS para Concursos 2017Natiele Crepaldi100% (1)
- Técnicas anestésicas para maxila e mandíbulaDocumento21 páginasTécnicas anestésicas para maxila e mandíbulaCHARLES DOS SANTOS NEVES100% (2)
- Perio teste conhecimentoDocumento4 páginasPerio teste conhecimentoArmando JúniorAinda não há avaliações
- Apostila Módulo02Documento49 páginasApostila Módulo02Armando JúniorAinda não há avaliações
- Cirurgia Odontológica APSDocumento8 páginasCirurgia Odontológica APSArmando JúniorAinda não há avaliações
- Apostila Módulo03Documento40 páginasApostila Módulo03Armando JúniorAinda não há avaliações
- Manual de Odontologia HospitalarDocumento32 páginasManual de Odontologia HospitalarArmando JúniorAinda não há avaliações
- Princípios e estrutura do SUSDocumento22 páginasPrincípios e estrutura do SUSArmando Júnior100% (10)
- Caderno Do DentistaDocumento49 páginasCaderno Do DentistaClaudiony Azevêdo100% (2)
- Preparos e restaurações odontológicasDocumento106 páginasPreparos e restaurações odontológicasmarc10oceanbluestarAinda não há avaliações
- Apostila EndodontiaDocumento101 páginasApostila EndodontiaArmando JúniorAinda não há avaliações
- Cariologia BásicaDocumento20 páginasCariologia BásicaArmando JúniorAinda não há avaliações
- O Posto de Trabalho Odontologico - ErgonomiaDocumento10 páginasO Posto de Trabalho Odontologico - ErgonomiaArmando JúniorAinda não há avaliações
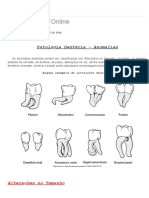
- Anomalias dentárias: microdontia, macrodontia e outras alteraçõesDocumento15 páginasAnomalias dentárias: microdontia, macrodontia e outras alteraçõesArmando JúniorAinda não há avaliações
- Manifestações orais em pacientes oncológicos tratados com quimioterapiaDocumento1 páginaManifestações orais em pacientes oncológicos tratados com quimioterapiaArmando JúniorAinda não há avaliações
- Acessibilida Na Usp de BauruDocumento13 páginasAcessibilida Na Usp de BauruArmando JúniorAinda não há avaliações
- Agenesias DentariasDocumento3 páginasAgenesias DentariasArmando JúniorAinda não há avaliações
- Imunologia - Sistema Imune InatoDocumento9 páginasImunologia - Sistema Imune InatoArmando JúniorAinda não há avaliações
- Farmacologia - Resumo CompletoDocumento95 páginasFarmacologia - Resumo CompletoCarol Pimenta100% (1)
- Estudo Dirigido - BioestatisticaDocumento3 páginasEstudo Dirigido - BioestatisticaArmando JúniorAinda não há avaliações
- Posições e movimentos da ATMDocumento9 páginasPosições e movimentos da ATMJoana MartinsAinda não há avaliações
- MEDRESUMOS 2016 - IMUNOLOGIA 08 - MHC e Apresentação Antigênica PDFDocumento8 páginasMEDRESUMOS 2016 - IMUNOLOGIA 08 - MHC e Apresentação Antigênica PDFArmando JúniorAinda não há avaliações
- O Segredo de Um Bom NegocioDocumento2 páginasO Segredo de Um Bom NegocioArmando JúniorAinda não há avaliações
- Órgãos Do Sistema ImuneDocumento5 páginasÓrgãos Do Sistema ImuneArmando JúniorAinda não há avaliações
- Mapa Conceitual - Reparo TecidualDocumento1 páginaMapa Conceitual - Reparo TecidualJOYCE FAZOLLO FERRAZAinda não há avaliações
- Casos ECG-TRAUMADocumento2 páginasCasos ECG-TRAUMACarol MarquesAinda não há avaliações
- Sistema Circulatório - Principais ConceitosDocumento2 páginasSistema Circulatório - Principais ConceitosAnacaby100% (9)
- Tabela Dose Maxima Diaria AinesDocumento1 páginaTabela Dose Maxima Diaria AinesSuzanna LimaAinda não há avaliações
- 19 - AnestésicosDocumento7 páginas19 - AnestésicosMarcos SoaresAinda não há avaliações
- Resumo PediatriaDocumento6 páginasResumo PediatriaalicelindaAinda não há avaliações
- DescriçãoDocumento81 páginasDescriçãoJonnathann MarvysAinda não há avaliações
- Aula Exsanguineotransfusão Mari 2023Documento26 páginasAula Exsanguineotransfusão Mari 2023Karolyne DuarteAinda não há avaliações
- Patologia Associada Ao Trigo.. Alergia IgE e Não IgE Mediada, Doença Celíaca...Documento17 páginasPatologia Associada Ao Trigo.. Alergia IgE e Não IgE Mediada, Doença Celíaca...Quéren VillasAinda não há avaliações
- Manual Da TacDocumento8 páginasManual Da TacKleber E Elaine FrancoAinda não há avaliações
- Roteiro padronizado de anamnese e exame físicoDocumento8 páginasRoteiro padronizado de anamnese e exame físicoFernanda FariasAinda não há avaliações
- Artrite EnteropáticaDocumento8 páginasArtrite EnteropáticaAndressa FerreiraAinda não há avaliações
- v95 Dermatoscopia Digital Metodo Complementar No Diagnostico Da EscabioseDocumento3 páginasv95 Dermatoscopia Digital Metodo Complementar No Diagnostico Da EscabioseNatalia TroncosoAinda não há avaliações
- FthnjufsccvcddghsDocumento3 páginasFthnjufsccvcddghsErickJunShinjoAinda não há avaliações
- Fisiologia Exercicio Fixação 03Documento6 páginasFisiologia Exercicio Fixação 03Fábio TelesAinda não há avaliações
- Pseudociese 1Documento9 páginasPseudociese 1Gianlucca AguiarAinda não há avaliações
- TCC Completo Daiane Soares - 2021Documento25 páginasTCC Completo Daiane Soares - 2021Silvia Regina Santos DE OliveiraAinda não há avaliações
- Boletim BibliográficoDocumento14 páginasBoletim Bibliográficocristovaoantonio antonioAinda não há avaliações
- Anemias - Ferropriva, B12, Aplásica e FalciformeDocumento15 páginasAnemias - Ferropriva, B12, Aplásica e FalciformeLeticia CostaAinda não há avaliações
- Choque: causas, sinais e tratamentoDocumento5 páginasChoque: causas, sinais e tratamentofrazao13Ainda não há avaliações
- Plano de parto otimizado paraDocumento2 páginasPlano de parto otimizado paraSaionara BennemannAinda não há avaliações
- Febre na Criança - Sinais, Medição e TratamentoDocumento31 páginasFebre na Criança - Sinais, Medição e TratamentoJoão CaladoAinda não há avaliações
- Características do medicamento veterinário EficurDocumento24 páginasCaracterísticas do medicamento veterinário EficurMarta RatoAinda não há avaliações
- Teorias Do Envelhecimento FinalDocumento9 páginasTeorias Do Envelhecimento FinalRoger SanglardAinda não há avaliações
- Questionário sobre dor pós-cirúrgica abdominal em idososDocumento6 páginasQuestionário sobre dor pós-cirúrgica abdominal em idososRoseni CabralAinda não há avaliações
- Adrenérgicos e AntiadrenérgicosDocumento23 páginasAdrenérgicos e AntiadrenérgicosLidmc100% (7)
- Relatório de Estágio - AMANDA SILVADocumento16 páginasRelatório de Estágio - AMANDA SILVAVall ReisAinda não há avaliações
- Mutarelli Propedêutica Neurológica PDFDocumento94 páginasMutarelli Propedêutica Neurológica PDFAna CarolinaAinda não há avaliações
- ELEMENTOS DA FARMACOLOGIA PriscilaDocumento79 páginasELEMENTOS DA FARMACOLOGIA PriscilaLucas Serra MartinsAinda não há avaliações
- Osteoporose: Doença Óssea que Aumenta Risco de FraturasDocumento3 páginasOsteoporose: Doença Óssea que Aumenta Risco de FraturasLuisa Loreto FernandesAinda não há avaliações