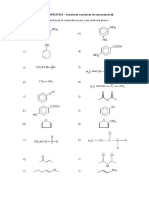
Você também pode gostar
- Resumo - Caso 02 (RNA)Documento8 páginasResumo - Caso 02 (RNA)jailsonteixeira.medeirosAinda não há avaliações
- Genética Médica para Médicos e Estudantes de Medicina: OncologiaNo EverandGenética Médica para Médicos e Estudantes de Medicina: OncologiaAinda não há avaliações
- Aula 2 Organização Gênica e TranscriçãoDocumento7 páginasAula 2 Organização Gênica e TranscriçãoRenaly AraujoAinda não há avaliações
- Revista de Ciência Elementar: Catarina MoreiraDocumento3 páginasRevista de Ciência Elementar: Catarina Moreiradldomingas341Ainda não há avaliações
- Biologia - Informação GenéticaDocumento6 páginasBiologia - Informação GenéticaJoão AzevedoAinda não há avaliações
- Tema 7 - Transcrição Do Dna - Versão Final PDFDocumento50 páginasTema 7 - Transcrição Do Dna - Versão Final PDFPedro SilvaAinda não há avaliações
- Genética geral: transcrição do DNA e classes de RNADocumento42 páginasGenética geral: transcrição do DNA e classes de RNAPedro SilvaAinda não há avaliações
- Síntese ProteicaDocumento27 páginasSíntese ProteicaInês PipaAinda não há avaliações
- DNA e Síntese Protéica - Passei DiretoDocumento6 páginasDNA e Síntese Protéica - Passei DiretoManuela ManuAinda não há avaliações
- Expressão Gênica PDFDocumento41 páginasExpressão Gênica PDFNayane AntunesAinda não há avaliações
- Processamento de RNA mensageiro em eucariotasDocumento3 páginasProcessamento de RNA mensageiro em eucariotasairton barrosAinda não há avaliações
- DNA, transcrição e síntese proteicaDocumento6 páginasDNA, transcrição e síntese proteicaMarcela TimossiAinda não há avaliações
- DNA replicação e transcriçãoDocumento10 páginasDNA replicação e transcriçãofernandalinsAinda não há avaliações
- 7 - Sintese ProtéicaDocumento10 páginas7 - Sintese ProtéicaCinthia MoraesAinda não há avaliações
- Resumos 1 Bio ST TesteDocumento15 páginasResumos 1 Bio ST TesteXavier VideiraAinda não há avaliações
- GENÉTICA HUMANA (Questões Prova)Documento6 páginasGENÉTICA HUMANA (Questões Prova)Marcos Felipe TrevisanAinda não há avaliações
- Transcricao e Processamento de RNA PDFDocumento43 páginasTranscricao e Processamento de RNA PDFcmartinsAinda não há avaliações
- TRANSCRIÇÃO E EXPRESSÃO GÊNICA: OS DIFERENTES TIPOS DE RNADocumento9 páginasTRANSCRIÇÃO E EXPRESSÃO GÊNICA: OS DIFERENTES TIPOS DE RNAInara NogueiraAinda não há avaliações
- Síntese de ProteinasDocumento3 páginasSíntese de ProteinasMarta OliveiraAinda não há avaliações
- Ácidos Nucléicos e Código GenéticoDocumento7 páginasÁcidos Nucléicos e Código GenéticoBordo NinaAinda não há avaliações
- Síntese Proteica: Do DNA aos AminoácidosDocumento7 páginasSíntese Proteica: Do DNA aos AminoácidosSusana MoraisAinda não há avaliações
- Trabalho Genética 1 Período MedicinaDocumento1 páginaTrabalho Genética 1 Período MedicinabrunaAinda não há avaliações
- DNA Transcrição RNAm ProcessamentoDocumento14 páginasDNA Transcrição RNAm ProcessamentoEsm KátyaAinda não há avaliações
- Síntese ProteicaDocumento1 páginaSíntese ProteicaSamantha LetíciaAinda não há avaliações
- Introdução Biologia MolecularDocumento56 páginasIntrodução Biologia MolecularGlenda Villarroel SilvaAinda não há avaliações
- Biologia celular: transcrição e traduçãoDocumento30 páginasBiologia celular: transcrição e traduçãoKamila LopesAinda não há avaliações
- Exercícios Do Texto 4 em Anexo Partes A, B e C.Documento7 páginasExercícios Do Texto 4 em Anexo Partes A, B e C.cadsus susAinda não há avaliações
- Sintese Proteica, Codigo Genetico, MutaçõesDocumento4 páginasSintese Proteica, Codigo Genetico, MutaçõesMargarida SaraivaAinda não há avaliações
- Processo de transcrição do DNA em RNADocumento3 páginasProcesso de transcrição do DNA em RNACassius RodrigoAinda não há avaliações
- A Replicação e Transcrição Do Dna - Ext. 2023Documento45 páginasA Replicação e Transcrição Do Dna - Ext. 2023Carol Bakos100% (1)
- RNA polimerase e transcrição genéticaDocumento7 páginasRNA polimerase e transcrição genéticaBruno CordeiroAinda não há avaliações
- Biologia Celular - Transcrição e TraduçãoDocumento57 páginasBiologia Celular - Transcrição e TraduçãomargaablourencoAinda não há avaliações
- Síntese de proteínas: transcrição e traduçãoDocumento3 páginasSíntese de proteínas: transcrição e traduçãoMarta OliveiraAinda não há avaliações
- Bioquímica Área IVDocumento9 páginasBioquímica Área IVNilza UFRGSAinda não há avaliações
- Código genético e a síntese de proteínasDocumento6 páginasCódigo genético e a síntese de proteínasfabiofapoAinda não há avaliações
- Resumo de Biologia 11 (Meu)Documento40 páginasResumo de Biologia 11 (Meu)Afonso CorreiaAinda não há avaliações
- Dna e RnaDocumento15 páginasDna e Rnamj.repinaldoAinda não há avaliações
- Genética básica para veterináriosDocumento12 páginasGenética básica para veterináriosDácia ArrudaAinda não há avaliações
- Seminário Citologia - TranscriçãoDocumento25 páginasSeminário Citologia - TranscriçãoCláudio Filho GomidesAinda não há avaliações
- Artigo Encontro 3 Pré - Diferença Entre mRNA - tRNA - rRNADocumento3 páginasArtigo Encontro 3 Pré - Diferença Entre mRNA - tRNA - rRNAtalitaAinda não há avaliações
- TranscriçãoDocumento2 páginasTranscriçãomônica conceiçãoAinda não há avaliações
- DNA e RNADocumento39 páginasDNA e RNAabelhudo gamerAinda não há avaliações
- Estrutura Do Gene e TranscriçãoDocumento5 páginasEstrutura Do Gene e TranscriçãoCaroline S. de SouzaAinda não há avaliações
- Resumo Estrutura Do DNA, Replicação, Transcrição, Tradução e MutaçãoDocumento15 páginasResumo Estrutura Do DNA, Replicação, Transcrição, Tradução e MutaçãombtbotinhaAinda não há avaliações
- DNA e síntese proteicaDocumento38 páginasDNA e síntese proteicaSoraia MartinsAinda não há avaliações
- Aula 6 Material de Apoio Transcrição e TraduçãoDocumento10 páginasAula 6 Material de Apoio Transcrição e TraduçãoRenaly AraujoAinda não há avaliações
- RESUMO - AV2 GenéticaDocumento8 páginasRESUMO - AV2 GenéticaMarina VieiraAinda não há avaliações
- Bruno. Transcrissao e Processamento de DnaDocumento3 páginasBruno. Transcrissao e Processamento de DnaBruno alberto AndreAinda não há avaliações
- Processamento de RNADocumento8 páginasProcessamento de RNALetícia CaeliAinda não há avaliações
- A Síntese de ProteínasDocumento24 páginasA Síntese de ProteínasThiaya Freitas MattosAinda não há avaliações
- Biologia Molecular da TranscriçãoDocumento38 páginasBiologia Molecular da TranscriçãoSamuel Castelo BrancoAinda não há avaliações
- Bruno. Transcrissao e ProcessamentoDocumento10 páginasBruno. Transcrissao e ProcessamentoBruno alberto AndreAinda não há avaliações
- Resumo de BiologiaDocumento2 páginasResumo de BiologiaLucia DuarteAinda não há avaliações
- Mapa mental da transcrição e traduçãoDocumento3 páginasMapa mental da transcrição e traduçãolais cerezolliAinda não há avaliações
- Ácidos Nucleicos - DNA e RNADocumento3 páginasÁcidos Nucleicos - DNA e RNAIvy MenezesAinda não há avaliações
- Resumos Completos Da Materia Biologia 11º para Exame Nacional 11 AnoDocumento17 páginasResumos Completos Da Materia Biologia 11º para Exame Nacional 11 AnoTomás RodriguesAinda não há avaliações
- TranscriçãoDocumento1 páginaTranscriçãomayara.luz2Ainda não há avaliações
- RNA MensageiroDocumento2 páginasRNA MensageiroSergio Miguel da SilvaAinda não há avaliações
- DNA e RNA: estrutura e funçãoDocumento7 páginasDNA e RNA: estrutura e funçãoLívia CottaAinda não há avaliações
- Observação Da VibraçãoDocumento1 páginaObservação Da VibraçãoMiriãAinda não há avaliações
- Replicação Do DnaDocumento44 páginasReplicação Do DnaMiriãAinda não há avaliações
- Genética Módulo 1Documento4 páginasGenética Módulo 1MiriãAinda não há avaliações
- Sistema Ocular SensorialDocumento4 páginasSistema Ocular SensorialMiriãAinda não há avaliações
- Sistema Visual MotorDocumento2 páginasSistema Visual MotorMiriãAinda não há avaliações
- As bases neurais da linguagem e o papel do HDDocumento25 páginasAs bases neurais da linguagem e o papel do HDMiriãAinda não há avaliações
- Filtração GlomerularDocumento3 páginasFiltração GlomerularMiriãAinda não há avaliações
- TARJABRANCAdocDocumento3 páginasTARJABRANCAdocMiriãAinda não há avaliações
- Processamento SensorialDocumento8 páginasProcessamento SensorialMiriãAinda não há avaliações
- Biofísica Eduardo A.C Garcia-2Documento382 páginasBiofísica Eduardo A.C Garcia-2MiriãAinda não há avaliações
- Chomsky - Teoria InatistaDocumento5 páginasChomsky - Teoria InatistaLuzia OliveiraAinda não há avaliações
- Biologia Geologia v1Documento16 páginasBiologia Geologia v1Lara RibeiroAinda não há avaliações
- FQ - Equilíbrio Químico - Revisão 2019Documento21 páginasFQ - Equilíbrio Químico - Revisão 2019Caio LimmaAinda não há avaliações
- Revisão - ENEM-SEMANA DDocumento25 páginasRevisão - ENEM-SEMANA DJedaAinda não há avaliações
- Propriedades periódicas e formação de iõesDocumento31 páginasPropriedades periódicas e formação de iõesAluno Ana Carolina Silva CabralAinda não há avaliações
- Atividade 1 - Seg - Segurança Do Trabalho Industrial - 51-2023Documento2 páginasAtividade 1 - Seg - Segurança Do Trabalho Industrial - 51-2023Cavalini Assessoria AcadêmicaAinda não há avaliações
- Enunciados Exemplos - AdsorçãoDocumento30 páginasEnunciados Exemplos - AdsorçãoFelipe Castro100% (1)
- Moléculas, Fórmulas Químicas e ReaçõesDocumento11 páginasMoléculas, Fórmulas Químicas e ReaçõesanitadiogoAinda não há avaliações
- Resumo - Revisões 10º Ano BiomoléculasDocumento14 páginasResumo - Revisões 10º Ano BiomoléculasBarbara CostaAinda não há avaliações
- Ficha CS 1 1Documento3 páginasFicha CS 1 1Alexander OliveiraAinda não há avaliações
- Artigo - Corrosão 2 Unid PDFDocumento6 páginasArtigo - Corrosão 2 Unid PDFNicoly CoelhoAinda não há avaliações
- Certificado de Pós-Graduação Quimica Geral e Organica Genivaldo FaveniDocumento2 páginasCertificado de Pós-Graduação Quimica Geral e Organica Genivaldo FaveniGustavoAinda não há avaliações
- Ctic5 em Ficha ÁguaDocumento4 páginasCtic5 em Ficha ÁguaSofia DiasAinda não há avaliações
- Tabela Grifo OficialDocumento12 páginasTabela Grifo OficialGRIFO LABORATORY100% (1)
- AULA 3 - Peeling, Hidratação e AcnesDocumento74 páginasAULA 3 - Peeling, Hidratação e AcnesThayane Diniz100% (1)
- Exercícios Sobre Nomenclatura de Hidrocarbonetos - Toda Matéria - SoladoDocumento10 páginasExercícios Sobre Nomenclatura de Hidrocarbonetos - Toda Matéria - Soladodarlan danramAinda não há avaliações
- Protac Nortox AdDocumento6 páginasProtac Nortox AdAparecido Renato de Deus dos Santos SantosAinda não há avaliações
- Exercicios de Estru Ressonancia 2 PDFDocumento5 páginasExercicios de Estru Ressonancia 2 PDFGui LhermeAinda não há avaliações
- Kps e solubilidade de saisDocumento10 páginasKps e solubilidade de saisRodrigo Gabriel BastosAinda não há avaliações
- Ficha Tecnica HT 10000Documento1 páginaFicha Tecnica HT 10000Saldao GoianoAinda não há avaliações
- Catalogo Vimaplas Web 180312183639Documento76 páginasCatalogo Vimaplas Web 180312183639LM_S_SAinda não há avaliações
- Estudo Dos Elementos Ferro (Fe), Cobalto (Co) e Cobre (Cu)Documento33 páginasEstudo Dos Elementos Ferro (Fe), Cobalto (Co) e Cobre (Cu)MARIA SERPAAinda não há avaliações
- Memorial Descritivo Hidrossanitário R06Documento6 páginasMemorial Descritivo Hidrossanitário R06Secretaria de Planejamento Governador Celso RamosAinda não há avaliações
- k22 SecurDocumento14 páginask22 SecuraparecidoAinda não há avaliações
- Gestão de riscos e inspeções em ambientes explosivosDocumento6 páginasGestão de riscos e inspeções em ambientes explosivosLuiz0% (1)
- Aula Destilacao MulticomponentesDocumento18 páginasAula Destilacao MulticomponentesThais OliveiraAinda não há avaliações
- Processamento do alho negro para maximizar antioxidantesDocumento132 páginasProcessamento do alho negro para maximizar antioxidantesiris jungesAinda não há avaliações
- Aula Geometria Molecular e PolaridadeDocumento31 páginasAula Geometria Molecular e PolaridadeGabriela ParroAinda não há avaliações
- Efeito da adição de nitrogênio na microestrutura e propriedades mecânicas do ADI austemperadoDocumento1 páginaEfeito da adição de nitrogênio na microestrutura e propriedades mecânicas do ADI austemperadoLeonardo PereiraAinda não há avaliações
- Manual do Usuário Kit GNV LandirenzoDocumento44 páginasManual do Usuário Kit GNV LandirenzoJunior MacielAinda não há avaliações
- Manual de Operação - 908EDocumento168 páginasManual de Operação - 908EMicaella SerafimAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Fundamentos Em Cirurgia OralNo EverandFundamentos Em Cirurgia OralNota: 5 de 5 estrelas5/5 (1)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Educação ambiental: A formação do sujeito ecológicoNo EverandEducação ambiental: A formação do sujeito ecológicoAinda não há avaliações
- Parasitologia 1: Helmintos de Interesse MédicoNo EverandParasitologia 1: Helmintos de Interesse MédicoAinda não há avaliações
- Serviços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesNo EverandServiços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesAinda não há avaliações
- Aprender mais rápido: Habilidades de estudo, memória e neurociênciaNo EverandAprender mais rápido: Habilidades de estudo, memória e neurociênciaNota: 4 de 5 estrelas4/5 (5)
- A clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNo EverandA clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNota: 3.5 de 5 estrelas3.5/5 (3)
- Fisiologia Cardiorrespiratória Ensino SuperiorNo EverandFisiologia Cardiorrespiratória Ensino SuperiorAinda não há avaliações
- Espécies nativas para piscicultura no Brasil: 3ª edição revista, atualizada e ampliadaNo EverandEspécies nativas para piscicultura no Brasil: 3ª edição revista, atualizada e ampliadaNota: 5 de 5 estrelas5/5 (2)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- Comportamento Animal: Uma Introdução aos Métodos e à Ecologia ComportamentalNo EverandComportamento Animal: Uma Introdução aos Métodos e à Ecologia ComportamentalNota: 3 de 5 estrelas3/5 (1)
- Concentração: Torne-se mais consciente e economize tempoNo EverandConcentração: Torne-se mais consciente e economize tempoNota: 4.5 de 5 estrelas4.5/5 (3)