Você também pode gostar
- Apostila Nutrição AnimalDocumento35 páginasApostila Nutrição AnimalRosinalldo Santos100% (2)
- Pregando o Evangelho PDFDocumento1 páginaPregando o Evangelho PDFMariaClaraBravanelAinda não há avaliações
- Sugestão e Estratégias para Os 50 Dias de Jejum e Oração RESUMIDO PDFDocumento2 páginasSugestão e Estratégias para Os 50 Dias de Jejum e Oração RESUMIDO PDFCristiano Mascarenhas da SilvaAinda não há avaliações
- Texto 2 - Trivelato e SilvaDocumento8 páginasTexto 2 - Trivelato e SilvaGabriel Nunes da SilvaAinda não há avaliações
- Apostila Dermaplaning LiliDocumento5 páginasApostila Dermaplaning LiliNeliane PimentaAinda não há avaliações
- Pas Saude 6566 - Ft2Documento3 páginasPas Saude 6566 - Ft2Joao Guedes100% (2)
- Aula 3Documento24 páginasAula 3benjaminbrazalvesAinda não há avaliações
- Leite de AlpisteDocumento1 páginaLeite de AlpisteMarlon Sebastião ChagasAinda não há avaliações
- Anais Jorp2012Documento207 páginasAnais Jorp2012Ana Paula GodoiAinda não há avaliações
- Livres de Satanás - Rev. Jeremias PereiraDocumento43 páginasLivres de Satanás - Rev. Jeremias PereiraadionirbAinda não há avaliações
- A Oração Na Vida Do CrenteDocumento4 páginasA Oração Na Vida Do CrenteDomingos MachadoAinda não há avaliações
- 02 - A Estrada - Revelações e Pensamentos GnósticosDocumento3 páginas02 - A Estrada - Revelações e Pensamentos GnósticosEpaminondas VimalakirtiAinda não há avaliações
- Epafras e o Dia de Oração - Joel BeekeDocumento3 páginasEpafras e o Dia de Oração - Joel BeekeMuriel Munique100% (1)
- Exegese de Atos 6.1-7Documento5 páginasExegese de Atos 6.1-7Alfredo DA Cruz Barbosa JúniorAinda não há avaliações
- EliasDocumento19 páginasEliasSiberio Melo100% (1)
- Carta Às 7 Igrejas - FiladélfiaDocumento7 páginasCarta Às 7 Igrejas - FiladélfiajackaugustAinda não há avaliações
- Apostasia Esfriamento Da Fé Tem Tomado Conta Da TerraDocumento6 páginasApostasia Esfriamento Da Fé Tem Tomado Conta Da TerraRaquel FragosoAinda não há avaliações
- O Princípio Da SantidadeDocumento1 páginaO Princípio Da SantidadeNéia Do CarlosAinda não há avaliações
- Apostila Panorama de Profetas Maiores e MenoresDocumento24 páginasApostila Panorama de Profetas Maiores e MenoresJoão MedeirosAinda não há avaliações
- Aula 19 - Exegese Sêmio-Discursiva de Marcos 3,13-19Documento4 páginasAula 19 - Exegese Sêmio-Discursiva de Marcos 3,13-19tiprofiAinda não há avaliações
- Introducao Codigo Apocalipse HanegraaffDocumento10 páginasIntroducao Codigo Apocalipse HanegraaffPaulo GondimAinda não há avaliações
- A Doutrina Do Espírito SantoDocumento28 páginasA Doutrina Do Espírito SantoGilberto LimaAinda não há avaliações
- Seitas e HeresiasDocumento27 páginasSeitas e HeresiasHugoPérisséAinda não há avaliações
- Manual Metodista Livre - Conbras 2007Documento139 páginasManual Metodista Livre - Conbras 2007j.ildo100% (4)
- As Sete Taças Do ApocalipseDocumento4 páginasAs Sete Taças Do ApocalipsePrLeandro SilvaAinda não há avaliações
- Tipologia PDFDocumento13 páginasTipologia PDFDaniele MarinhoAinda não há avaliações
- Apstila de Escatologia - AocDocumento21 páginasApstila de Escatologia - AocadoniasbgAinda não há avaliações
- A Queda de LúciferDocumento13 páginasA Queda de LúciferbsantosrjAinda não há avaliações
- Lição A Grande TribulaçãoDocumento10 páginasLição A Grande TribulaçãoPriscilia FreitasAinda não há avaliações
- A Mulher Adultera No Evangelho de JoãoDocumento16 páginasA Mulher Adultera No Evangelho de JoãoBelchior TobiasAinda não há avaliações
- Em Busca de Um Renovo EspiritualDocumento4 páginasEm Busca de Um Renovo EspiritualLIVIAAinda não há avaliações
- 2 - As Duas Naturezas Do CrenteDocumento5 páginas2 - As Duas Naturezas Do Crentefabiojs37100% (1)
- Seminário de EscatologiaDocumento12 páginasSeminário de EscatologiaThamar Soares100% (1)
- GalatasDocumento8 páginasGalatasJunior Marques100% (1)
- Soteriologia BentesDocumento59 páginasSoteriologia BentespastorbentesgoelAinda não há avaliações
- Aula 3 Pentateuco - Ninrode À Babilônia (Contexto Religioso No A.t.)Documento15 páginasAula 3 Pentateuco - Ninrode À Babilônia (Contexto Religioso No A.t.)camis100% (1)
- Teologia Do Novo TestamentoDocumento55 páginasTeologia Do Novo TestamentoDaniel MartinsAinda não há avaliações
- Hebraicoe Feedicao 1Documento18 páginasHebraicoe Feedicao 1Francisco SoutoAinda não há avaliações
- 1b. Artigo - Culto e Espiritualidade PentecostalDocumento13 páginas1b. Artigo - Culto e Espiritualidade PentecostalAndré Luís da Rosa100% (1)
- A Parabola Do SemeadorDocumento5 páginasA Parabola Do SemeadorJelvan Mendes100% (1)
- O Sinal de JonasDocumento35 páginasO Sinal de JonasFamarteAinda não há avaliações
- Tanque de BetesdaDocumento4 páginasTanque de BetesdaEduardo PiresAinda não há avaliações
- Curso de Doutrina Biblica Rev Antonio AlDocumento79 páginasCurso de Doutrina Biblica Rev Antonio AlMariosergio Gouvea100% (1)
- Anõnimos Que Fazem A DiferençaDocumento2 páginasAnõnimos Que Fazem A DiferençaHercilio CelinoAinda não há avaliações
- Avivamento No Antigo TestamentoDocumento6 páginasAvivamento No Antigo TestamentoAna SuelyAinda não há avaliações
- Vida DevocionalDocumento21 páginasVida DevocionalAlberto Santos100% (1)
- A PÓS-MODERNIDADE, UM DESAFIO À PREGAÇÃO-Isaltino GomesDocumento18 páginasA PÓS-MODERNIDADE, UM DESAFIO À PREGAÇÃO-Isaltino GomesFransuarlem BarbosaAinda não há avaliações
- Atos 8 (20032021)Documento15 páginasAtos 8 (20032021)jefferso_cv100% (2)
- O Pai Dos LuzesDocumento12 páginasO Pai Dos LuzesSilvia Cristina100% (2)
- Comentário Bíblico - Provérbios A CantaresNo EverandComentário Bíblico - Provérbios A CantaresAinda não há avaliações
- Cristo MaravilhosoDocumento0 páginaCristo MaravilhosoJúnior MedeirosAinda não há avaliações
- Jesus Cristo e A Ordem de MelquisedequeDocumento2 páginasJesus Cristo e A Ordem de MelquisedequeDiego R GomesAinda não há avaliações
- LeviticoDocumento12 páginasLeviticonuro2010Ainda não há avaliações
- Lição 01Documento23 páginasLição 01Luiz RibeiroAinda não há avaliações
- Memórias de Charles FinneyDocumento326 páginasMemórias de Charles Finneyricl100% (1)
- Arminianismoecalvinismo 120420102843 Phpapp02Documento2 páginasArminianismoecalvinismo 120420102843 Phpapp02Rose Meire Oliveira BarrosAinda não há avaliações
- Aula 9 Pastoreio Mútuo e DiscipuladoDocumento50 páginasAula 9 Pastoreio Mútuo e DiscipuladoClayton VelozoAinda não há avaliações
- O Profeta AmósDocumento10 páginasO Profeta AmósIngrid AlmeidaAinda não há avaliações
- Surpreendidos Pela GraçaDocumento8 páginasSurpreendidos Pela GraçaReginaldo Adriana SousaAinda não há avaliações
- Por Que A Igreja Adventista Do Setimo Dia Eh Uma SeitaDocumento6 páginasPor Que A Igreja Adventista Do Setimo Dia Eh Uma SeitaRodrigo LucianaAinda não há avaliações
- Como Ler e Citar Uma Referência BíblicaDocumento3 páginasComo Ler e Citar Uma Referência BíblicaMarcos RamosAinda não há avaliações
- Escatologia e ApocalipseDocumento2 páginasEscatologia e ApocalipseSione VieiraAinda não há avaliações
- As Marcas de Uma Igreja Que Abalou o MundoDocumento2 páginasAs Marcas de Uma Igreja Que Abalou o MundoDavid BarrosAinda não há avaliações
- Sermão - Lucas 24.13-35 - Os Discípulos No Caminho de EmaúsDocumento3 páginasSermão - Lucas 24.13-35 - Os Discípulos No Caminho de EmaúsArmando AntunesAinda não há avaliações
- Ebd - Livro de Ezequiel - CompartilharDocumento7 páginasEbd - Livro de Ezequiel - Compartilharjazz ghostAinda não há avaliações
- Edital Pub Usp 2018 2019 PDFDocumento9 páginasEdital Pub Usp 2018 2019 PDFGabriel Nunes da SilvaAinda não há avaliações
- AtomicaMolecularScript PDFDocumento181 páginasAtomicaMolecularScript PDFGabriel Nunes da SilvaAinda não há avaliações
- Controle Endócrino Do Metabolismo EnergéticoDocumento9 páginasControle Endócrino Do Metabolismo EnergéticoGabriel Nunes da SilvaAinda não há avaliações
- Procariotos e EucariotosDocumento11 páginasProcariotos e EucariotosGabriel Nunes da SilvaAinda não há avaliações
- Aula 2Documento23 páginasAula 2Gabriel Nunes da SilvaAinda não há avaliações
- Determinação de Vitamina CDocumento6 páginasDeterminação de Vitamina CMirela RosaAinda não há avaliações
- ImunohematologiaDocumento34 páginasImunohematologiaMarta Oliveira100% (2)
- Sinais e Sintomas Clínico NutricionaisDocumento11 páginasSinais e Sintomas Clínico NutricionaisPatyLeandroAinda não há avaliações
- AnelideosDocumento9 páginasAnelideosJair Ferreira LimaAinda não há avaliações
- Brinde Aula Chave Mestra Do EmagrecimentoDocumento13 páginasBrinde Aula Chave Mestra Do EmagrecimentoJoão André RodriguesAinda não há avaliações
- Massagem Com Pedras QuentesDocumento48 páginasMassagem Com Pedras QuentesKatia Regina Petersen100% (5)
- Anamnese PadrãoDocumento8 páginasAnamnese PadrãoAlisson DouradoAinda não há avaliações
- (2013) Hipertensao Gestacao PPT UFPR PDFDocumento82 páginas(2013) Hipertensao Gestacao PPT UFPR PDFGabriel SampaioAinda não há avaliações
- Abdome Agudo PDFDocumento75 páginasAbdome Agudo PDFkeityAinda não há avaliações
- Chondrichthyes GeralDocumento104 páginasChondrichthyes GeralCelso Cardeal Jr.67% (3)
- Aula 2 - Farmacocinética e Farmacodinâmica Av1Documento42 páginasAula 2 - Farmacocinética e Farmacodinâmica Av1Marcus Vinicius CardosoAinda não há avaliações
- 4 - Clostridiose AviáriaDocumento48 páginas4 - Clostridiose AviáriaFuligemzAinda não há avaliações
- ApresentaçãoDocumento349 páginasApresentaçãoGabriel100% (1)
- Desenvolvimento Do Sistema NervosoDocumento57 páginasDesenvolvimento Do Sistema NervosoFabbio BaldoinoAinda não há avaliações
- Vet - Protocolo - ZoletilDocumento8 páginasVet - Protocolo - ZoletilLeandro MendesAinda não há avaliações
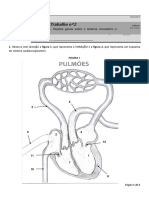
- Trabalho de Anatomia Do Trato Respiratorio Do Homem ISCEDDocumento10 páginasTrabalho de Anatomia Do Trato Respiratorio Do Homem ISCEDmussa nassordino nassordinoAinda não há avaliações
- AULA 3.2 - ART GERAL - 12.03.14 - Receptores Toll-LikeDocumento8 páginasAULA 3.2 - ART GERAL - 12.03.14 - Receptores Toll-LikeTiago OliveiraAinda não há avaliações
- Farmacologia Aplicada À Enfermagem Administração de Quimioterápicos e NPTDocumento40 páginasFarmacologia Aplicada À Enfermagem Administração de Quimioterápicos e NPTJanice MooreAinda não há avaliações
- Teste Global - Obtenção de EnergiaDocumento10 páginasTeste Global - Obtenção de EnergiaJoana GinjasAinda não há avaliações
- Miologia Dos Membros InferioresE.F AVA 2019Documento22 páginasMiologia Dos Membros InferioresE.F AVA 2019Claudio Sergio da CostaAinda não há avaliações
- Chifrei o Meu MaridoDocumento6 páginasChifrei o Meu MaridoSergius Magnus AndradeAinda não há avaliações
- Avaliandocincias4anosistemarespiratrio 150416150607 Conversion Gate02Documento2 páginasAvaliandocincias4anosistemarespiratrio 150416150607 Conversion Gate02Mary C R SouzaAinda não há avaliações
- Exercícios de BioestatísticaDocumento3 páginasExercícios de BioestatísticaRafael Henrique Rodrigues AlvesAinda não há avaliações
- Espermograma - Análise Do SêmemDocumento12 páginasEspermograma - Análise Do SêmemMarilene LopesAinda não há avaliações