Escolar Documentos
Profissional Documentos
Cultura Documentos
Bento Tratado de Implante Coclear 978-65-5572-084-6 Cap 04
Enviado por
Sabrina FigueiredoTítulo original
Direitos autorais
Formatos disponíveis
Compartilhar este documento
Compartilhar ou incorporar documento
Você considera este documento útil?
Este conteúdo é inapropriado?
Denunciar este documentoDireitos autorais:
Formatos disponíveis
Bento Tratado de Implante Coclear 978-65-5572-084-6 Cap 04
Enviado por
Sabrina FigueiredoDireitos autorais:
Formatos disponíveis
CAPÍTULO 4
FISIOLOGIA DO SISTEMA AUDITIVO
Mauricio Kurc
INTRODUÇÃO do meato acústico e a do pavilhão geram um ganho de 15 a 20 dB
O papel do sistema auditivo é converter o estímulo mecânico do na membrana timpânica em relação à entrada do canal. Acima e
som em sensação auditiva no córtex cerebral. Pode ser dividido, abaixo de 3,5 kHz o ganho é progressivamente menor. O ganho de
do ponto de vista funcional e anatômico, em periférico e central. intensidade do som nessa frequência é uma das razões da perda au-
O aparelho auditivo periférico engloba uma unidade que recebe o ditiva induzida por ruído ocorrer inicialmente na região de 4 kHz.
som do meio ambiente, a orelha externa; um sistema de condução
da energia mecânica, a orelha média; e um sistema ultraespeciali- Audição Direcional
zado de amplificação e codificação dessa energia, a cóclea, locali- A audição direcional, ou seja, a capacidade de detectar a origem da
zada na orelha interna. fonte sonora no plano horizontal, é explicada principalmente pe-
O aparelho auditivo diferencia-se dos demais órgãos envolvi- las diferenças na intensidade e no tempo de chegada do som nas
dos com os sentidos. O órgão sensorial auditivo é classificado como duas orelhas.
um mecanorreceptor por converter a energia mecânica do som em Como as orelhas estão localizadas em lados opostos da cabe-
impulsos elétricos no nervo auditivo, enquanto as células sensoriais ça, o tempo de chegada do estímulo sonoro é diferente para cada
da retina dos olhos (os cones e bastonetes) são fotorreceptores por orelha, dependendo da distância de cada uma delas em relação à
transformarem a energia eletromagnética da luz em impulsos elé- fonte sonora. Quanto mais longe da fonte, maior o tempo para che-
tricos. Já os órgãos olfatórios e gustatórios são classificados como gada do estímulo.
quimiorreceptores. Em relação à intensidade, a cabeça age como um obstáculo para
O aparelho auditivo central, formado pelo nervo coclear, pelos o som que chega à orelha oposta, diminuindo a intensidade do som
núcleos das vias auditivas do tronco cerebral e mesencéfalo, córtex que chega nessa orelha (efeito sombra) (Fig. 4-1). Por outro lado,
auditivo temporal e vias subcorticais associativas, é responsável pelo a intensidade do som que chega à orelha próxima à fonte sonora
processamento dos impulsos elétricos gerados pelo sensor perifé- é aumentada pelo efeito concha exercido pela cabeça. No sistema
rico e proporciona a sensação de tonalidade e intensidade, além da nervoso central, essas diferenças são processadas, e o cérebro de-
atribuição de significado (cognição) a esses estímulos. termina a localização da fonte sonora.
É importante lembrar que, além da função auditiva, a orelha O tempo de chegada do som independe da frequência. Já a in-
tensidade do som que chega às orelhas depende da frequência do
interna também participa da manutenção do equilíbrio corporal
som como também da posição da fonte sonora em relação ao plano
por meio do aparelho vestibular. Esse órgão é responsável pela
horizontal. A cabeça age como um atenuador somente se o compri-
percepção da posição da cabeça no espaço, também através de
mento da onda sonora for menor que a largura da cabeça. Assim,
estímulos mecânicos, ou seja, os movimentos angulares e linea-
com frequências maiores que 2 kHz, o efeito sombra chega a causar
res da cabeça.
uma diferença interaural de até 15 dB. Com frequências menores,
Neste capítulo focaremos o funcionamento do sistema auditivo
nas quais o comprimento de onda é maior que o comprimento da
periférico. Quando não está funcionando adequadamente, este pode,
cabeça, o efeito sombra não é tão significativo. No plano vertical, o
em alguns casos, ser substituído pelo implante coclear.
mecanismo de localização da fonte sonora não é bem compreendi-
do, mas provavelmente também está relacionado às propriedades
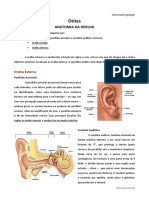
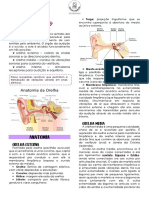
ORELHA EXTERNA acústicas do pavilhão, com suas elevações, saliências e sua forma
A função da orelha externa, composta pelo pavilhão auricular e pelo em concha, e do meato acústico externo.
meato acústico externo, é receber os sons do ambiente e conduzi-
-los para a orelha média. ORELHA MÉDIA
Os sons medidos no meato acústico externo são consideravel-
Composta pela membrana timpânica, pelos ossículos (martelo, bi-
mente diferentes dos sons medidos num campo sonoro. gorna e estribo) e pelos músculos tensor do tímpano e estapédio, a
A razão para essa diferença tem sido analisada por duas pers- orelha média é responsável por transmitir o som da orelha externa
pectivas. Uma é a influência das ressonâncias que a orelha externa para a orelha interna. A tuba auditiva é um canal que comunica a
faz no som que chega à membrana timpânica. A outra é a influência orelha média à rinofaringe e tem a função de manter a pressão do
da orelha externa na percepção de direcionalidade, o que ajuda na ar no interior da orelha média próximo à pressão do ar no meio
localização da fonte sonora. ambiente, com o objetivo de otimizar a condução do som.
Quando o estímulo acústico alcança a membrana timpânica
Ressonância do Meato Acústico Externo pode chegar à orelha interna através de três formas: por condução
O meato acústico externo é simplesmente um tubo aberto de um óssea, alcançando diretamente a cóclea sem passar pelos ossícu-
lado e fechado do outro, funcionando como uma cavidade de res- los; por difusão pelo ar da cavidade timpânica e através da cadeia
sonância. A frequência de ressonância é determinada pelo compri- ossicular. Obviamente, esse último é o meio mais efetivo, como ve-
mento do tubo. O meato acústico externo de um adulto tem, em remos a seguir.
média, 2,5 cm. Para esse comprimento, a frequência de ressonância A vibração da membrana timpânica desencadeada pelo som
é de, aproximadamente, 3,5 kHz. Nessa frequência, a ressonância é transmitida para a cadeia ossicular. A vibração dos ossículos
39
© 2021. Thieme. All rights reserved.
40 PARTE II SISTEMA AUDITIVO
em média, de 55 mm2 e a área da janela oval, de 3,2 mm2. Assim, a
pressão do som agindo sobre a membrana timpânica é concentra-
da na área muito menor da janela oval, gerando amplificação, pois
Pressão (p) é igual a Força (F) dividida pela Área (A) (p = F/A). Como
a pressão exercida na janela oval é obtida pela razão das duas áre-
as (55 mm2/3,2 mm2), a pressão resultante na platina do estribo é
17 vezes maior que na membrana timpânica, correspondendo a um
ganho de aproximadamente 25 dB.
O segundo mecanismo é o de alavanca, que ocorre entre os
ossículos da orelha média. A extensão do manúbrio do martelo é
1,3 vezes maior que a extensão do processo longo da bigorna (Fig.
4-2, mm/rlb). Assim, uma pequena força aplicada ao manúbrio do
martelo resulta num ganho de força no ramo longo da bigorna.
O terceiro mecanismo deve-se à forma côncava da membrana
timpânica que se estica quando se movimenta, fazendo com que o
martelo se mova com aproximadamente o dobro da força.
Teoricamente, se as estruturas da orelha média funcionassem
idealmente, o resultado de todos esses mecanismos combinados
seria 17 × 1,3 × 2 = 44,2, ou seja, um aumento de 44 vezes na pres-
são que chega ao interior da cóclea, correspondendo a um ganho de
33 dB. Entretanto, o poder de transformação da orelha média huma-
na não é perfeito, e alguma energia sonora é refletida pela membra-
na timpânica, como revelam os resultados de diferentes investigado-
res, que mostram um ganho da orelha média na faixa de 25 a 30 dB.
O ganho obtido pela orelha média também varia conforme a
frequência do estímulo. Merchant et al. chegaram a valores de ga-
nho de aproximadamente 20 dB entre 250 e 500 Hz e um ganho
máximo de 25 dB em 1.000 Hz.1 Até 200 Hz a eficiência da orelha
média humana fica cerca de 10 dB abaixo do ideal. Sua maior efici-
ência ocorre ao redor de 1 kHz. Nessa frequência, fica apenas 3 dB
abaixo de um transformador de impedâncias ideal.
Impedância Acústica
A impedância acústica é uma medida da resistência que a membrana
timpânica apresenta ao ser colocada em movimento pelo som. Pode
fornecer informações importantes sobre o mecanismo de condução
Fig. 4-1. Esquema ilustrando o efeito sombra. A cabeça age como um do estímulo sonoro para a cóclea e, portanto, é muito utilizada na
obstáculo para o som que chega à orelha oposta, diminuindo a intensidade do investigação das afecções da orelha média.
som que chega nessa orelha (efeito sombra). Na orelha mais próxima à fonte A timpanometria é uma forma de determinar a impedância da
sonora, a intensidade do som é aumentada pelo efeito concha que a cabeça orelha média, e a variação da impedância acústica é utilizada para
exerce. A cabeça e o pavilhão auditivo agem como atenuadores somente identificar contrações do músculo estapédio, com o intuito de es-
se o comprimento da onda sonora for menor que a largura da cabeça. Esse
tudar os reflexos acústicos.
efeito é mais importante nas frequências maiores que 2 kHz. Com frequências
menores, nas quais o comprimento de onda é maior que o comprimento da A medida da impedância acústica tem uma alta reprodutibili-
cabeça, o efeito sombra não é tão significativo. dade no mesmo indivíduo, mas pode variar muito entre indivíduos
pelas diferenças da membrana timpânica, principalmente das pro-
priedades da sua porção flácida.
movimenta a platina do estribo, e essa vibração é transmitida aos A membrana timpânica, os ossículos, os músculos, o ar da cavi-
líquidos cocleares, às estruturas da cóclea e à janela redonda. É jus- dade timpânica e a cóclea contribuem para a impedância da orelha
tamente a presença da janela redonda e de sua membrana compla- média. Contudo, experimentos em gatos e coelhos, em que a arti-
cente que permite o movimento da platina do estribo. Quando o culação íncudoestapediana é desarticulada, mostraram que a im-
estribo se movimenta para dentro da cóclea, a membrana da janela pedância acústica da orelha é diminuída para valores bem baixos,
redonda move-se para fora. Esse acoplamento reflete a ocorrência sugerindo que o principal componente da impedância acústica da
de uma onda hidromecânica no interior da cóclea. orelha é a cóclea.
A Orelha Média Como um Transformador de Músculos da Orelha Média
Impedâncias Existem dois músculos estriados dentro da orelha média: o múscu-
A orelha média funciona como um transformador de impedâncias lo tensor do tímpano e o estapédio. O músculo tensor do tímpano
porque transmite a energia mecânica do som do meio aéreo, de está ligado ao martelo, é inervado pelo nervo trigêmeo e, quando
baixa impedância (baixo atrito), para o interior líquido da cóclea, se contrai, puxa a membrana timpânica medialmente. O músculo
que é um meio de alta impedância (alto atrito). Se não houvesse estapédio conecta-se ao estribo e é inervado pelo nervo estapedia-
esse efeito transformador da orelha média e o som fosse conduzido no, ramo do nervo facial. Quando se contrai, de forma reflexa, com
diretamente do ar para a janela oval, menos de 1% da energia me- sons a partir de 80 dB, puxa o estribo perpendicularmente ao mo-
cânica chegaria ao interior da cóclea. vimento de pistão que esse ossículo exerce na janela oval. O reflexo
Esse processo de compensação da energia é realizado pela ore- do estapédio ocorre bilateralmente, mesmo com estímulo unilateral.
lha média através de três mecanismos. O principal deles é a dife- São os dois menores músculos do corpo e, embora sua contra-
rença entre a área da membrana timpânica e a área da platina do ção inegavelmente altere a transmissão do som através da orelha
estribo na janela oval, ou mais precisamente, a razão entre essas média para a cóclea, sua exata função e a magnitude da sua ação
áreas (Fig. 4-2, a razão Amt/Ae). A área da membrana timpânica é, não estão completamente esclarecidas.
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 41
Fig. 4-2. (a) Esquema do sistema tímpano-ossicular mostrando as diferenças na área e no padrão de vibração dos ossículos. Ae: Área do estribo; Amt: área da
membrana timpânica; rlb: comprimento do ramo longo da bigorna; mm: comprimento do manúbrio do martelo. (b) Fotografia da orelha média de cadáver
humano sob visualização endoscópica. As paredes do meato acústico externo foram ressecadas. Mm: manúbrio do martelo; rlb: ramo longo da bigorna;
tme: tendão do musculo estapédio; ta: tuba auditiva; e: estribo; m: martelo; b: bigorna; llm: ligamento lateral do martelo; lsm: ligamento superior do martelo.
(Imagem b cortesia de João Paulo Valente).
Os reflexos dos músculos da orelha média são lentos. Sua latên- ORELHA INTERNA – CÓCLEA
cia diminui conforme o aumento na intensidade do estímulo, de 150 O papel da cóclea é converter a energia mecânica do som em impul-
até 35 mseg, no máximo da sua contração. Além disso, a contração
sos elétricos, o que é denominado de transdução mecanoelétrica.
do músculo vai aumentando até se consolidar totalmente, podendo
Esse processo é complexo, mas fascinante, pois envolve mecanismos
demorar até 500 mseg para se contrair maximamente. Em razão
acústicos, mecânicos, elétricos, químicos e neurais.
dessa demora na contração dos músculos, o reflexo não tem efei-
to em mudanças rápidas das intensidades dos sons a que estamos Os movimentos vibratórios do sistema tímpano-ossicular de-
expostos. Mudanças rápidas na intensidade são transmitidas para sencadeados pelo som geram pressão na janela oval e energia hidro-
a cóclea com pouca ou nenhuma atenuação. Outra característica mecânica no interior fluido da cóclea. Acredita-se, mas também se
desse reflexo é que ele funciona mais rápido e mais intensamente questiona, que essa energia desencadeie uma onda que viaja atra-
para sons abaixo de 1.000 Hz. Acima dessa frequência, o efeito ate- vés da escala vestibular, atravessa o helicotrema e alcança a escala
nuante da contração do músculo estapédio é pequeno e não fun- timpânica em direção à janela redonda (Fig. 4-3).2
ciona como um mecanismo regulatório efetivo.
O fato de que o reflexo da orelha média influencia mais as baixas
frequências do que as médias e altas frequências pode ser impor-
tante para reduzir os efeitos do mascaramento que sons de baixas
frequências exercem sobre os de alta frequência.
Em pacientes com paralisia do nervo facial, com paralisia total
do músculo estapédio unilateral, ocorre maior mudança temporal
do limiar auditivo na orelha afetada após a exposição a sons de bai-
xa frequência do que na orelha com reflexo do estapédio normal.
A inteligibilidade da fala acima de 90 dB também fica prejudicada
nas pessoas sem o reflexo do estapédio, quando comparadas com
pessoas com reflexo normal.
Os reflexos da orelha média são ativados pela fala da própria
pessoa num grau maior que seria esperado pela intensidade do
som que realmente chega à orelha interna. E o reflexo pode ser de-
tectado antes mesmo que a fala tenha começado. Essa ação pode
ser importante para preservar a audição durante a fala ou como Fig. 4-3. Esquema da via de transmissão do som da membrana timpânica
efeito antimascaramento, prevenindo que componentes de baixa à cóclea. Os estímulos sonoros incidem na membrana timpânica (mt), e
frequência da fala da própria pessoa mascarem outros sons de fre- os movimentos vibratórios, denotados por setas, são transmitidos através
quências mais altas. dos três ossículos da orelha média: martelo (m), bigorna (b) e estribo (e).
O músculo tensor do tímpano não está envolvido na atenuação A platina do estribo comporta-se como um pistão na janela oval e inicia
dos sons. Diferentemente de alguns animais, como gatos, coelhos e ondas de pressão nos fluidos cocleares, pondo em vibração a membrana
basilar. A pressão é aliviada na janela redonda (jr). A cóclea neste esquema
cobaia, no ser humano o músculo tensor do tímpano não responde
está retificada, mas, na verdade, tem a forma de uma concha de caracol e
reflexamente a sons intensos. Sua contração puxa o manúbrio do fica incrustada na porção petrosa do osso temporal. É subdividida em três
martelo, causando um movimento da membrana timpânica para compartimentos contendo líquidos denominados perilinfa ou endolinfa. Os
dentro. Ele contrai durante a deglutição, quando a tuba auditiva é compartimentos externos são conectados no helicotrema. O comprimento
aberta, auxiliando a troca de ar na cavidade timpânica. total da cóclea é de 35 mm nos seres humanos. (Modificado de Fettiplace.)2
© 2021. Thieme. All rights reserved.
42 PARTE II SISTEMA AUDITIVO
O movimento diferencial entre a lâmina basilar e a membrana Pelo menos durante os estágios iniciais de seu desenvolvimento,
tectória, desencadeado por essa onda, resulta no estímulo dos me- todos os tipos de estereocílios apresentam a mesma conformação
canorreceptores sensoriais auditivos, as células ciliadas do órgão de duas ou mais fileiras de estereocílios e um único cinocílio. O ci-
espiral de Corti. nocílio está presente nos estereocílios da maioria dos órgãos, mas
As células ciliadas da cóclea estão dispostas em uma fileira única desaparece dos estereocílios da cóclea dos mamíferos durante os
de células ciliadas internas (CCI) e em três fileiras de células ciliadas estágios iniciais do desenvolvimento pós-natal e antes da matura-
externas (CCE), dispostas de forma tonotópica ao longo do ducto ção da audição. Os estereocílios são microvilos modificados, densa-
coclear (Fig. 4-4a). Sons de alta frequência ativam as células cilia- mente preenchidos por filamentos de actina (Fig. 4-5), e o cinocílio,
das da base da cóclea e sons de baixa frequência ativam as células quando presente, possui o característico arranjo dos cílios móveis
ciliadas do ápice, com um gradiente entre elas. Essa organização (9 duplas de microtúbulos em torno de um par de microtúbulos
tonotópica é mantida em toda a via auditiva, desde o 1o neurônio centrais).
no gânglio espiral até os centros corticais superiores no lobo tem- Somente os estereocílios das células ciliadas externas fazem
poral, passando pelos vários núcleos auditivos no tronco cerebral. contato com a membrana tectória e são estimulados quando ocorre
Cada célula ciliada contém em sua superfície apical um feixe o movimento diferencial entre a membrana basilar e a membrana
de 50 a 150 estereocílios dispostos em um padrão de escada, com tectória (Fig. 4-4d,e). Os estereocílios das células ciliadas internas
várias fileiras em alturas decrescentes (Fig. 4-4b). O feixe de estere- não fazem contato direto com a membrana tectória.
ocílios é a estrutura mecanossensível que permite a conversão das
vibrações induzidas pelo estímulo sonoro em respostas elétricas, Transdução Mecanoelétrica
retransmitidas ao sistema nervoso central (SNC) com precisão e A transdução mecanoelétrica se inicia com a deflexão do feixe de
fidelidade surpreendentes. estereocílios desencadeada pelas oscilações da membrana basilar
Os feixes de estereocílios aparecem numa variedade de tama- (Fig. 4-6). Resultante dessa deflexão, ocorre a abertura de canais
nhos em diferentes órgãos e espécies e têm forma e função sob me- catiônicos não seletivos, ou seja, canais que permitem a passagem
dida relativos aos parâmetros do estímulo a que são submetidos. de qualquer íon positivo.3
Fig. 4-4. (a) Eletromicrografias de varredura da membrana reticular contendo 3 fileiras de células ciliadas externas e 1 fileira de células ciliadas internas.
A membrana reticular é completada pelas porções apicais dos processos falangianos das células de suporte e pelas porções apicais das células dos pilares.
(b,c) Eletromicrografias de varredura da região apical de células ciliadas externas de cobaia, nas quais se observam os feixes de estereocílios em forma de W.
(d) Visão inferior da membrana tectória onde se visualizam as impressões dos estereocílios mais longos das células ciliadas externas. (e) Coroas de fixação que
conectam os estereocílios mais longos das células ciliadas externas à membrana tectória.
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 43
o potencial intracelular das células ciliadas é, em média, -70 mV nas
células ciliadas externas (Fig. 4-7).5
É no momento da abertura mecânica dos canais de transdução
e do resultante influxo de corrente elétrica que ocorre a transdução
mecanoelétrica, ou seja, a conversão do estímulo mecânico do som
em informação útil para o receptor auditivo e para o SNC.
A despolarização celular ativa funções diferentes nas células ci-
liadas. Na célula ciliada interna, abre canais de cálcio voltagem-de-
pendentes, gerando influxo de cálcio e, consequentemente, libera-
ção de glutamato pelas vesículas sinápticas e ativação de receptores
ionotrópicos de glutamato AMPA das terminações pós-sinápticas
das fibras aferentes do nervo auditivo. Na célula ciliada externa, o
ciclo despolarização/hiperpolarização desencadeia alterações da
voltagem da membrana plasmática que resultam em respostas me-
cânicas de alongamento e encurtamento dessas células.
Quando as células ciliadas externas se encurtam, a membrana
basilar aproxima-se da lâmina reticular e reduz o espaço subtecto-
rial, gerando um fluxo radial de endolinfa que resulta no estímulo
dos estereocílios das células ciliadas internas. Essas células são con-
sideradas os verdadeiros receptores auditivos, uma vez que recebem
90 a 95% das aferências que saem da cóclea pelo nervo auditivo. Em
contrapartida, as células ciliadas externas, apesar de serem três
vezes mais frequentes na cóclea (Fig. 4-4), recebem apenas 5% da
inervação aferente.
Estrutura do Feixe de Estereocílios
O feixe de estereocílios deflete-se como um todo, em razão de um
complexo de conexões existentes entre cada estereocílio, e pivo-
ta na sua base, junto à sua inserção na superfície apical da célula
Fig. 4-5. A estrutura de actina no interior dos estereocílios confere rigidez ao ciliada. Nota-se que as porções basais dos estereocílios são mais
feixe estereociliar. (a) Eletromicrografia de varredura do feixe estereociliar de afiladas em relação ao seu corpo, o que contribui para esse pivota-
cobaia. Essa especialização da célula ciliada é mantida ereta pela citoesqueleto mento (Fig. 4-8a).6
interno de actina e pelas conexões entre os estereocílios. (b) Eletromicrografia As conexões entre os estereocílios descritas são as conexões
de transmissão mostrando o denso empacotamento de actina no interior do tornozelo (ankle links), as conexões de haste (shaft links), as co-
dos estereocílios. Observe também a conexão de ponta unindo o estereocílio
nexões superiores (top links) e as conexões de ponta (tiplinks) (Fig.
maior à ponta de um estereocílio menor e as respectivas densidades superior
4-8b).7 No órgão vestibular existe uma 5ª conexão chamada cinoci-
e inferior, sugerindo forte concentração de proteínas. (Modificado de
Fettiplace.)2 (c) Eletromicrografia de transmissão de uma réplica de platina e liar, que acopla o cinocílio ao estereocílio imediatamente adjacente.
carbono de um feixe de estereocílios de células ciliadas da ampola de cobaia, As conexões do tornozelo localizam-se logo acima do ponto
preparada pela técnica de congelamento rápido e freeze-etching, em que se onde cada estereocílio se insere na placa cuticular. Essas conexões
visualizam as conexões laterais entre os estereocílios. (d) Maior aumento da provavelmente são formadas por duas grandes proteínas USH2A e
figura anterior mostrando o denso citoesqueleto de filamentos de actina no ADGRV1. Ambas são produtos de genes responsáveis pela síndro-
interior do estereocílio. me de Usher tipo 2, uma doença que causa surdez congênita e ce-
gueira progressiva.
A direção do movimento do feixe estereociliar influencia a aber- As conexões de haste são vistas como finos filamentos ou como
uma malha entre estereocílios adjacentes. A proteína PTPRQ, pro-
tura do canal de transdução mecanoelétrica. Deflexão na direção
duto do lócus responsável pela DFNB84, é o componente provável
do estereocílio mais alto abre os canais, enquanto a deflexão em
dessas conexões.
direção aos estereocílios mais baixos os fecha. Deflexões laterais
As conexões superiores variam em estrutura conforme a espécie
não têm efeito (Fig. 4-6).4 e o tipo de célula ciliada. Nas células ciliadas externas dos mamífe-
Em virtude da altíssima concentração de potássio na endolin- ros, as conexões superiores possuem uma estrutura parecida com
fa, líquido extracelular que banha a superfície apical das células um zíper: filamentos finos projetam-se das membranas dos este-
ciliadas, a abertura do canal produz um influxo maciço de cargas reocílios opostos e condensam-se dentro de uma densidade extra-
positivas na forma de potássio (K+), gerando uma corrente de recep- celular central (Fig. 4-8c).8 Embora nas células ciliadas internas as
ção. Uma pequena parte dessa corrente é levada por íons de cálcio conexões superiores conectem as regiões laterais dos estereocílios
(Ca+2), também presente na endolinfa, mas em menor concentração. tanto entre fileiras como ao longo das fileiras de estereocílios, como
Na verdade, o canal de transdução das células ciliadas é mais ocorre nas células ciliadas externas, uma densidade central não é
permeável ao cálcio, mas é o potássio que leva a maior parte da observada e pode estar ausente nesse tipo de célula.
corrente de transdução por sua concentração ser muito maior no Em camundongos, a estereocilina (STRC), um produto do lócus
fluido que banha a região apical da célula ciliada, a endolinfa. A es- DFNB16, está associada às conexões superiores das células ciliadas
externas, bem como às coroas de fixação (Fig. 4-4), que acoplam
tria vascular ativamente injeta potássio (contra esse gradiente de
as pontas dos estereocílios mais altos à membrana tectorial sobre-
concentração) no interior do ducto coclear.
jacente. Como a STRC não é detectada nos estereocílios das células
O feixe é extremamente sensível ao estímulo mecânico. Defle-
ciliadas internas, provavelmente contribui para a densidade central,
xões menores que o diâmetro de um átomo são suficientes para que até agora só foi observada nas conexões superiores das células
iniciar a transdução mecânica. Uma das razões é o elevado potencial ciliadas externas.
elétrico da endolinfa na ordem de +80 mV, denominado de poten- As conexões de ponta, que conectam os estereocílios ao longo do
cial endococlear, ausente no vestíbulo e nos canais semicirculares eixo de sua sensibilidade mecânica, são fundamentais para a trans-
e mantido pela alta concentração de potássio. Isso gera uma força dução mecanoelétrica.9-11 Acredita-se que a conexão de ponta seja
elétrica de 150 mV para a entrada de potássio e cálcio, uma vez que responsável pela abertura do canal de transdução mecanoelétrica.
© 2021. Thieme. All rights reserved.
44 PARTE II SISTEMA AUDITIVO
Fig. 4-6. A transdução mecanoelétrica se inicia com a deflexão do feixe de estereocílios desencadeada pelas oscilações da membrana basilar resultando na
abertura dos canais de transdução. O aparelho de transdução das células ciliadas está localizado no topo do feixe de cabelo e inclui uma conexão de ponta que
interconecta estereocílios adjacentes. A extremidade inferior da conexão de ponta parece que está conectada a dois canais mecanicamente sensíveis.
A extremidade superior termina na placa de inserção, local onde estão as moléculas de miosina Ic envolvidas na adaptação do processo de transdução. Quando
o estímulo mecânico deflete o feixe de estereocílios na direção positiva (diagrama à direita) em direção aos estereocílios mais longos, os estereocílios pivotam
nas suas inserções basais, mas permanecem em contato nas pontas. O movimento resultante entre os estereocílios contíguos aumenta a tensão na conexão da
ponta e aumenta a probabilidade de que os canais na inserção inferior sejam abertos. Uma corrente iônica, transportada predominantemente por potássio, mas
incluindo cálcio, entra então no estereocílio, despolariza a célula ciliada e desencadeia a liberação de glutamato nas sinapses das células ciliadas internas e ações
motoras nas células ciliadas externas. (Modificado de Hudspeth.)4
estereocílios.13 Após o estímulo, a fluorescência foi visualizada ape-
nas nas extremidades superiores dos estereocílios menores, estando
ausentes no estereocílio mais alto, demonstrando que a localização
precisa dos canais é na parte inferior da conexão de ponta e que não
existem canais de transdução nos estereocílios na fileira mais alta.
Os Canais de Transdução Mecanoelétrica São Abertos
Mecanicamente
Os sistemas sensoriais como audição, visão e olfato são bastante
sensíveis e capazes de detectar sinais muito pequenos. Assim, en-
Fig. 4-7. Potencial endococlear. O elevado potencial elétrico da endolinfa na quanto bastonetes fotorreceptores detectam um único fóton e recep-
ordem de +80 mV, denominado de potencial endococlear, é o responsável
tores olfatórios são sensíveis a moléculas individuais de odorantes,
pela extrema sensibilidade do complexo de transdução mecanoelétrica das
células ciliadas. A diferença de potencial entre a endolinfa e o interior negativo os mecanorreceptores da orelha, as células ciliadas, são capazes de
das células ciliadas gera uma força elétrica de 150 mV nas células ciliadas detectar, no limiar auditivo, movimentos de dimensões atômicas
externas e de 120 mV nas células ciliadas internas. Deflexões menores que o (0,3 nm) em seu feixe de estereocílios.
diâmetro de um átomo são suficientes para que ocorra a entrada de potássio Apesar de algumas similaridades, contudo, esses receptores
e cálcio na célula e se inicie a transdução mecanoelétrica. Esse potencial utilizam estratégias diferentes para recepção, transmissão e ampli-
é mantido pela estria vascular, que, ativamente e contra um gradiente de
ficação dos estímulos. Enquanto os receptores visuais, gustativos e
concentração, mantém a alta concentração de potássio na endolinfa da escala
média ou ducto coclear. EM: escala média; CCI: célula ciliada interna; CCE:
olfatórios utilizam cascatas bioquímicas intracelulares que resul-
célula ciliada externa. (Modificada de Dallos.)5 tam na abertura dos canais iônicos, as células sensoriais auditivas
realizam esse processo através da abertura mecânica e direta dos
canais de transdução.
Por causa desse papel crucial, atraíram muita atenção e provavel- Há vantagens na utilização de cascatas bioquímicas intracelu-
mente são as conexões mais bem caracterizadas do feixe de este- lares, pois estas possibilitam tanto a amplificação, por intermédio
reocílios. do recrutamento de mensageiros secundários, como mecanismos
A extremidade inferior da conexão de ponta está acoplada a um de retroalimentação dentro da via metabólica, permitindo o con-
ou dois canais iônicos. A deflexão do feixe de estereocílios em di- trole de ganho da operação, como adaptação e dessensibilização.
reção aos estereocílios mais altos aumenta a tensão na conexão de A utilização de um mecanismo de transdução direto, sem in-
ponta, que funciona como uma mola de portão, puxando o canal e tervenção de mensageiros secundários, tem como vantagem a
ocasionando a sua abertura, enquanto a deflexão em direção aos es- velocidade da resposta. As células ciliadas operam muito mais
tereocílios menores desencadeia o fechamento do canal (Fig. 4-6).12 rapidamente do que qualquer outro receptor sensorial, mais rá-
pido até que os próprios neurônios. Essa velocidade é necessária,
Localização do Canal de Transdução Mecanoelétrica por exemplo, na audição direcional, onde a orelha humana precisa
A localização exata do canal de transdução mecanoelétrica nas cé- distinguir diferenças inferiores a 20 µs na chegada do som em cada
lulas ciliadas foi motivo de debate ao longo da história da pesquisa orelha para identificar a fonte sonora. A velocidade de resposta
em audição. Beurg et al. (2009) utilizaram microscopia confocal também é importante para lidar com as frequências auditivas da
para observar alterações rápidas de fluorescência, que refletiam ordem de 20.000 Hz, ou seja, o canal de transdução deve funcionar
a entrada de cálcio durante a estimulação mecânica de feixes de 20 mil vezes por segundo. No caso dos animais ecolocalizadores,
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 45
Fig. 4-8. Estrutura e conexões do feixe de estereocílios. (a) Eletromicrografia de transmissão de uma sessão longitudinal da
região apical de uma célula ciliada do sáculo de peixe dourado. Mostra a base afilada dos estereocílios contendo uma densa
concentração de filamentos de actina onde eles se inserem na placa cuticular.6 (b) Ilustração esquemática de uma seção
longitudinal através de três estereocílios localizados em filas adjacentes, conectados por conexões de ponta e outras mais. Os
quatro tipos conhecidos de conexões são mostrados, bem como as respectivas proteínas constituintes dessas conexões. Em
vermelho, as perdas auditivas genéticas correspondentes.7 (c, d) Eletromicrografia de transmissão de células ciliadas externas
de rato mostrando as conexões superiores dos estereocílios. Observe o condesamento central da conexão superior, como um
zíper (CDH23, caderina 23; PCDH15, protocaderina 15; USH1e USH2, mutações Usher tipo 1 e Usher tipo 2, USH2A, usherina;
ADGRV1, Adhesion G protein-coupled receptor V1; PTPRQ, protein-tyrosine phosphatase, receptor-type Q). (Imagens cortesia
de Richard Goodyear.)8
como baleias, golfinhos e morcegos, as frequências são da ordem
de 100.000 a 150.000 Hz.
Após a deflexão do feixe estereociliar, o canal de transdução
mecanoelétrica abre-se em questão de microssegundos, menos de
50 µs na cóclea de ratos, velocidade que exclui a possibilidade de
mensageiros secundários.12,14
ADAPTAÇÃO DO CANAL DE TRANSDUÇÃO
Imediatamente após a deflexão dos estereocílios, o canal de trans-
dução se fecha para interromper a entrada de correntes na célula
e terminar o estímulo. Esse mecanismo é chamado de adaptação
do canal de transdução. Dois mecanismos de adaptação foram des-
critos: um rápido, que ocorre em poucos milissegundos, e outro
mais lento, que ocorre após mais de 10 ms. A adaptação rápida
ocorre através da entrada de cálcio pelo canal de transdução, que
se liga diretamente a alguma parte desse canal e altera a tensão e
a probabilidade de o canal ficar aberto. O mecanismo de adapta- Fig. 4-9. Adaptação do canal de transdução. Diagrama representando os dois
ção lenta ocorre também pela entrada de íons cálcio, que ativam o modelos propostos para explicar a adaptação do canal de transdução: (a) um
deslizamento de moléculas de miosina ao longo dos feixes de ac- processo mais lento mediado por motor, que requer a redefinição da tensão
tina do estereocílio, reposicionando mais inferiormente a inserção da conexão da ponta feita pelo deslizamento das proteínas que compõem a
da conexão de ponta no estereocílio mais longo, reduzindo assim densidade superior da conexão da ponta ao longo dos filamentos de actina
o estiramento e determinando o fechamento do canal (Fig. 4-9).15 e (b) adaptação rápida e direta mediada pelo cálcio, que provavelmente
resulta da ligação de cálcio diretamente ao canal de transdução ou a proteínas
associadas ao canal, aumentando a probabilidade do seu fechamento.
Estrutura das Conexões de Ponta e a Transdução (Modificado de Holt e Corey.)15
Mecanoelétrica
A busca dos componentes principais da maquinaria de transdução
mecanoelétrica das células ciliadas vem ocorrendo há décadas. Nos ponta, como de proteínas que estão intrinsecamente ligadas à fun-
últimos anos, houve um progresso significativo na identificação das ção do canal de transdução mecanoelétrica. Esse progresso ocorreu,
moléculas que formam e regulam o canal de transdução sensorial, principalmente, através de estudos baseados na análise da função
com a identificação tanto das proteínas que formam as conexões de de genes ligados à perda auditiva.
© 2021. Thieme. All rights reserved.
46 PARTE II SISTEMA AUDITIVO
Fig. 4-10. Ultraestrutura das conexões de ponta (tiplinks). (a) Ilustração da provável estrutura da conexão de ponta, como um duplo filamento entrelaçado, que
une a ponta de um estereocílio ao seu vizinho mais alto. (b-d) Eletromicrografia de transmissão por freeze-etching de células ciliadas de cobaias. (b, c) Conexão
de ponta em maior resolução, onde claramente se visualizam os dois filamentos entrelaçados. (d) A conexão de ponta bifurca-se em dois pequenos ramos antes
de conectar um estereocílio mais alto. (e) Na outra extremidade, vários pequenos filamentos estendem-se da conexão de ponta ao estereocílio menor.16
Utilizando a técnica de freeze-fracture e deep-etching e visua- A estrutura que confere elasticidade, como a mola de portão
lizando a ultraestrutura desses filamentos por microscopia eletrô- (Fig. 4-11), ainda permanece controversa, mas pode incluir as pró-
nica, mostramos pela primeira vez que as conexões de ponta são prias caderinas das conexões de ponta, as moléculas ainda não iden-
compostas por um par de filamentos entrelaçados de forma heli- tificadas conectadas a essas caderinas ou alguma outra combinação.
coidal, que formam um filamento único com um diâmetro de 8 a Em cortes finos de estereocílios, visualizados por microscopia
10 nm (Fig. 4-10).16 Na extremidade superior, podem bifurcar-se, eletrônica, densidades submembranosas, conhecidas como densida-
e na extremidade inferior dois ou três ramos conectam a conexão
des superior e inferior, são observadas nos locais em que as partes
de ponta ao estereocílio mais curto.
superior e inferior das conexões de ponta ligam-se às membranas
O par de filamentos entrelaçados é formado por dois homodí-
dos estereocílios (Fig. 4-5).19,20
meros, um composto por duas moléculas de caderina 23 (CDH23) e
o outro por duas moléculas de protocaderina 15 (PCDH15), que se CDH23 e PCDH15 interagem com várias outras proteínas in-
associam para conectar um estereocílio mais curto ao seu próximo tracelulares. Essas proteínas formam um complexo que permite
vizinho mais alto. A CDH23 localiza-se na extremidade superior da a CDH23 interagir com o citoesqueleto de F-actina do estereocílio
conexão de ponta, e a PCDH15, na extremidade inferior (Fig. 4-11).17 e com fosfolipídios da membrana celular. Muitas dessas proteínas
Mutações nos genes que codificam CDH23 e PCDH15 são a causa são também codificadas por genes associados à síndrome de Usher
da síndrome de Usher tipo 1 (Figs. 4-8 e 4-11). (Figs. 4-8 e 4-11). O domínio citoplasmático da CDH23 interage
Na posição de repouso, o canal de transdução oscila entre aber- com as proteínas harmonina (USH1C), SANS (USH1G) e MYO7A
to e fechado aproximadamente mil vezes por segundo, mas está (USH1B). Na porção inferior da conexão de ponta, o domínio cito-
fechado na maior parte do tempo. plasmático da PCDH15 interage com, pelo menos, três proteínas:
Para que uma célula ciliada seja capaz de discriminar entre di- TMIE (proteína transmembrana da orelha interna), LHFPL5 (lipoma
ferentes intensidades sonoras, deflexões progressivamente maiores
HMIG fusion partner-like protein) e as proteínas transmembrana
do feixe de estereocílios aumentam a probabilidade de abertura
tipo-canal TMC1e TMC2.
dos canais gradativamente. Aumentos no estímulo aumentam a
A interação desse complexo da conexão de ponta com o cito-
tensão na conexão de ponta, ampliando a probabilidade de os ca-
nais abrirem. esqueleto é ainda modulada por várias outras proteínas. Na porção
No entanto, as conexões de ponta não têm característica elásti- superior, há um motor de miosina que controla a tensão da cone-
ca como se supunha.16 A conexão entre PCDH15 e CDH23 lembra a xão de ponta e a adaptação do canal de transdução mecanoelétrica
forma de um “aperto de mão”. Em simulações, essa região do aperto (MYO1C), como visto anteriormente, e na porção inferior, MYO15A,
de mão resiste bastante à tensão, alongando-se apenas alguns na- WHRN, ESPN-1 e ESPNL (duas proteínas associadas à actina) e a
nômetros, indicando ser uma estrutura bastante rígida.18 proteína ligadora de cálcio e integrina (CIB2) (Fig. 4-11).
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 47
como no endotélio, nas hemácias, na pele, em proprioceptores mus-
culares, receptores de insuflação pulmonar e barorreceptores na
aorta e no seio carotídeo.
Para que uma proteína seja considerada o canal iônico de me-
canotransdução, ela deve atender aos seguintes critérios: deve es-
tar localizada nas pontas dos estereocílios das células ciliadas e ser
necessária para a mecanotransdução nessas células, além de ser um
canal iônico mecanossensível formador de poros.
PIEZO2 também é expressa na porção apical das células ciliadas,
porém na base dos estereocílios, e não na ponta, onde se localiza o
canal de transdução. Sua abertura ocorre por pressão negativa na
superfície apical da célula e também por deslocamento do feixe
estereociliar em direção aos estereocílios menores (polaridade re-
versa) e independente dos tiplinks. A audição é apenas levemente
alterada pelo knockout de PIEZO2 e, portanto, não é essencial para
o funcionamento auditivo.23 Coletivamente, esses resultados afas-
taram a possibilidade de que PIEZO1 e 2 sejam as proteínas que
formam o canal de transdução. Talvez sejam apenas moléculas pre-
sentes no canal das células ciliadas em desenvolvimento inicial ou
após insultos em células ciliadas mais maduras.
Um corpo grande de evidências aponta para TMC1 e TMC2 como
as proteínas formadoras do poro do complexo do canal de transdu-
ção mecanoelétrica (Figs. 4-11 e 4-12).23-29
TMC1 e TMC2 são expressas por células ciliadas da orelha in-
terna24 e localizam-se nas fileiras de estereocílios mais baixos das
células ciliadas externas e das células ciliadas internas, na localiza-
ção correspondente aos canais de transdução (Figs. 4-11 e 4-12).27
Aminoácidos intracelulares de TMC1 e TMC2 interagem com a
PCDH15 na ponta dos estereocílios inferiores de ambas as células
ciliadas externas e células ciliadas internas.30 A mutação dominante
causadora da surdez de Beethoven em TMC1 altera a permeabili-
dade ao cálcio do canal de transdução mecanoelétrica. Além disso,
a corrente de transdução mecanoelétrica é totalmente suprimida
nos camundongos knockout duplo TMC1/TMC2, causando graves
déficits auditivos e de equilíbrio. No entanto, ela pode ser resgata-
da por vetores virais adenoassociados (AAVs) que expressam TMC1
ou TMC2.24,25,31 Conjuntamente, essas observações sugerem forte-
mente que TMC1/2 são componentes chaves do canal de transdu-
ção mecanoelétrica.
O perfil de expressão temporal difere para os dois genes e en-
Fig. 4-11. Esquema mostrando as interações moleculares entre a conexão de
ponta e os estereocílios. A conexão de ponta é formada por 2 filamentos que tre os órgãos auditivos e vestibulares. Na cóclea, TMC2 é expressa
se associam para conectar um estereocílio mais curto ao seu vizinho próximo nos primeiros dias de vida e depois desaparece, enquanto TMC1 é
mais alto. O par de filamentos entrelaçados superior é composto por duas expressa depois e mantida na idade adulta.24,32 Nos órgãos vestibu-
moléculas de caderina 23 (CDH23) e o filamento inferior por duas moléculas lares, os genes TMC1 e TMC2 permanecem expressos nas células
de protocaderina 15 (PCDH15). CDH23 e PCDH15 interagem com várias ciliadas vestibulares maduras.
outras proteínas intracelulares. O domínio citoplasmático da CDH23 interage
Além dessa diferença no perfil de expressão espaço-temporal,
com as proteínas harmonina (USH1C), SANS (USH1G) e MYO7A (USH1B). Na
porção inferior da conexão de ponta, o domínio citoplasmático da PCDH15 TMC1 e TMC2 desempenham papéis distintos. Nos canais forma-
interage com, pelo menos, três proteínas: TMIE (proteína transmembrana da dos apenas por TMC2, a seletividade para o cálcio é muito maior
orelha interna), LHFPL5 (lipoma HMIG fusion partner-like protein) e as proteínas que nos canais formados somente por TMC1. As correntes de trans-
transmembrana tipo-canal TMC1e TMC2. dução são maiores nas células ciliadas que expressam TMC2, por-
tanto as células que expressam TMC2 podem suportar um influxo
de cálcio significativamente maior. Nos órgãos vestibulares, onde
Estrutura do Canal de Transdução Mecanoelétrica não existe um grande potencial endococlear, a força motriz para
A identificação molecular do canal de transdução mecanoelétrica a entrada do cálcio na célula ciliada é menor, o que pode permi-
das células ciliadas, e de qualquer outra estrutura, tem sido um tir a expressão persistente de TMC2 na idade adulta. Na cóclea,
grande desafio ao longo da história pela pequena quantidade de o grande potencial endococlear deve exigir a mudança de desen-
tecido disponível no órgão auditivo, pelo pequeno número de ca- volvimento na expressão do TMC para canais com menor perme-
nais por célula, pela complexidade molecular desses canais e pela abilidade ao cálcio.33,34
dificuldade de expressar as subunidades do canal em outros siste- Como mencionado, a extremidade inferior da conexão de ponta
mas celulares clássicos para provar sua função.21,22 é composta por um homodímero de PCDH15,16,17,35 e cada monô-
As proteínas PIEZO1 e 2 foram as primeiras proteínas postula- mero de PCDH15 se ligaria a cada monômero TMC1 (Figs. 4-11 e
das como constituintes do canal de transdução mecanoelétrica das 4-12) para regular a abertura de um poro independentemente do
células ciliadas, por atenderem aos pré-requisitos necessários para outro.36 Não se sabe se a abertura desses dois poros é simultânea
tal e também por estarem presentes em outros mecanorreceptores, ou sequencial.
© 2021. Thieme. All rights reserved.
48 PARTE II SISTEMA AUDITIVO
Fig. 4-12. Modelo provável do aparelho de mecanotransdução das células ciliadas. (a) Micrografia
eletrônica de transmissão mostrando o estiramento pela conexão de ponta e formação de uma
tenda na membrana plasmática do estereocílio. (b) Esquema mostrando a localização do complexo
de mecanotransdução na ponta do estereocílio. (c) Um possível arranjo de 6 proteínas essenciais
para a transdução auditiva. TMC1 e PCDH15 são ambos dímeros, portanto pode-se supor que haja
uma relação um-para-um entre eles, com a PCDH15 abrindo o poro de uma subunidade TMC num
canal dimérico. Existem poucas informações sobre possíveis combinações para as outras proteínas
do complexo, nem se sabe quais são parte do complexo totalmente maduro e que são necessárias
apenas para a sua montagem durante o desenvolvimento. Compõem ainda o possível aparelho
LHFPL5, CIB2, TOMT e TMIE. (d) Estrutura terciária do TMC1 dimérico. As duas subunidades são
coloridas em azul e cinza. Dentro da subunidade azul, estão as hélices formadoras de poros. As setas
vermelhas mostram a via de permeação prevista em cada subunidade.
SELETIVIDADE DE FREQUÊNCIAS NA CÓCLEA região as fibras nervosas correspondentes seriam excitadas. Sons de
O entendimento de como a cóclea separa a informação auditiva num frequências diferentes envolveriam outros grupos de ressonadores,
espectro de frequências (discriminação de frequências) passou por em diferentes regiões e correspondendo a outras fibras nervosas.
quatro fases históricas distintas até chegarmos ao momento atual Helmholtz argumentou que, se gritássemos próximo às cordas de
de compreensão. um piano, algumas cordas ressonariam em correspondência às fre-
quências da voz. A lâmina basilar responderia similarmente, fazendo
assim uma análise do espectro de frequências do estímulo sonoro.
Primeira Fase (Elementos Ressonantes de Helmholtz)
Hermann Ludwig Ferdinand von Helmholtz37,38 foi o primeiro a su-
Segunda Fase (Onda Viajante)
gerir, em meados do século XIX, que seria na cóclea que ocorreria a
A primeira metade do século XX foi dominada pelos experimentos
discriminação de frequências, e não no cérebro. Influenciado pelas
do engenheiro húngaro-americano George von Békésy. Apesar de
então recentes descrições da anatomia do órgão espiral por Alfon-
considerar a teoria da ressonância de Helmholtz “a mais elegante
so Corti e outros anatomistas dos séculos XVIII e XIX, Helmholtz das teorias da audição”, ele não se contentou apenas com hipóteses
sugeriu que haveria dentro da cóclea elementos ressonantes inde- teóricas. Resolveu abrir a cóclea e observar o padrão vibratório da
pendentes e espacialmente ordenados para cada frequência. partição coclear durante o estímulo sonoro. Contratado pela com-
Apesar de não comprovar cientificamente suas hipóteses, nem panhia telefônica húngara para melhorar a qualidade das trans-
ter observado diretamente os movimentos dos fluidos cocleares, Hel- missões telefônicas, von Békésy resolveu primeiramente entender
mholtz postulou que fibras transversas da membrana basilar seriam como funcionava o sistema auditivo.
os minúsculos ressonadores, cada um sintonizado a uma frequência Ao estudar principalmente ossos temporais de cadáveres huma-
diferente. A sintonia variaria continuamente ao longo da membrana, e nos, descreveu a teoria da onda viajante, que forma grande parte do
o estímulo por um tom específico colocaria em vibração um pequeno que hoje compreendemos como ocorre a discriminação de frequên-
grupo de ressonadores numa determinada região, e somente nessa cias na cóclea.39 Utilizando ossos temporais isolados, von Békésy
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 49
fez aberturas na cápsula ótica, espalhou micropartículas reflexivas Além disso, os picos de vibração ocorriam numa região muito
na membrana vestibular intacta (membrana de Reissner), selou as extensa da cóclea, o que não explicava o alto grau de discriminação
aberturas com cimento e vidro e observou os movimentos dessa de frequências já observado em experimentos psicofísicos realiza-
membrana, em resposta a estímulos sonoros aplicados no meato dos pelo próprio von Békésy39,42 e nas respostas de núcleos auditi-
acústico externo. Assumiu que o padrão de vibração da membrana vos no SNC. Críticas também foram feitas quanto aos experimentos
vestibular assemelha-se ao padrão de vibração da lâmina basilar. terem sido realizados em cadáveres e, portanto, não refletirem o
Para poder observar esses movimentos ao microscópio ótico, usou funcionamento coclear in vivo.
iluminação estroboscópica, como se faz hoje para examinar as vi- von Békésy concluiu que a fraca resolução do processo de dis-
brações das pregas vocais. Observou que a vibração do estribo não criminação de frequências, observada em seus experimentos na
resulta em uma simples ressonância de componentes mecânicos cóclea post-mortem, seria uma consequência do alto grau de atrito
independentes, mas que, na verdade, as diferentes regiões da cóclea resultante da viscosidade do fluido presente no órgão espiral. Até
estão mecanicamente associadas. 1980, tanto ele como a maioria dos fisiologistas auditivos assumi-
Segundo suas conclusões, em resposta à vibração do estribo, uma ram que a audição dependeria de algum outro mecanismo para a
onda hidrodinâmica ocorre dentro da cóclea e propaga-se desde a discriminação fina de frequências. Aparentemente, as alterações pa-
base até o ápice, produzindo movimentos oscilatórios da lâmina basi- tológicas, e mesmo a morte, não teriam efeito sobre a onda viajante.
lar. Durante sua propagação, a onda viajante, como denominou, cres- A cóclea faria, então, uma análise espectral inicial e mais gros-
ce progressivamente em amplitude, atinge um pico e abruptamente seira do som. Um processo adicional de discriminação fina de fre-
decai (Fig. 4-13).40,41 O local de vibração máxima da onda varia com a quências, possivelmente uma análise neural dentro da cóclea ou no
frequência do estímulo, de tal forma que sons de alta frequência pro- sistema nervoso central, seria necessário para explicar essa discre-
duzem picos de vibração mais próximos à base da cóclea, enquanto pância entre a extraordinária sensibilidade e discriminação de fre-
tons de frequências baixas produzem picos mais próximos ao ápice. quências observadas nos experimentos psicofísicos e os movimen-
Essas características vibratórias da lâmina basilar seriam de- tos oscilatórios pouco sintonizados da lâmina basilar observados
terminadas, principalmente, por suas propriedades físicas, incluin- em cadáveres e vários outros animais.
do massa e rigidez, que variam gradualmente ao longo da cóclea. Quando foi demonstrado que as respostas das fibras do nervo
Na região basal, a lâmina basilar é mais fina e rígida, propiciando coclear tinham uma seletividade de frequências muito similar àque-
melhor vibração com estímulos de alta frequência, enquanto na la observada psicoacusticamente, ficou claro que a análise deveria
cúpula ela é mais espessa e flácida, facilitando a vibração com fre- ocorrer perifericamente, ou seja, no interior da cóclea.
quências mais baixas. Evans criou, então, o termo “segundo filtro”,43 que seria outro
O conceito revolucionário da onda viajante é certamente válido mecanismo que explicaria as diferenças na capacidade de discri-
ainda hoje e rendeu a von Békésy o prêmio Nobel de Fisiologia ou minação dos movimentos mecânicos observados na lâmina basilar
Medicina de 1961, o único prêmio Nobel agraciado à pesquisa do e as respostas altamente sintonizadas do nervo auditivo. Confor-
sistema auditivo até hoje. me suas palavras, “o mal sintonizado filtro da membrana basilar é
Entretanto, várias críticas foram feitas a esses experimentos. seguido por um segundo filtro, fisiologicamente vulnerável, linear,
Primeiro, para que os movimentos da lâmina basilar fossem visí- mas bem sintonizado, situado entre a membrana basilar e o não
veis, eram necessários estímulos sonoros de, pelo menos, 120 dB linear processo de geração de potencial de ação neural.”
NA, bem acima do limiar auditivo e que, na cóclea viva, causam le- Uma gama de esquemas foi criada para explicar o segundo fil-
são do órgão. Estímulos próximos ao limiar não proporcionavam tro, incluindo complexas redes neurais, mecanismos bioquímicos e
respostas vibratórias da lâmina basilar. diversos modelos matemáticos de processamento elétrico e mecâni-
A cóclea seria um sistema linear. O aumento da magnitude das co, porém a existência desse segundo filtro nunca foi demonstrada.
vibrações cocleares era linearmente proporcional ao aumento do
estímulo sonoro. No entanto, a natureza não é linear. Sistemas bio- Terceira Fase (Amplificador Coclear)
lógicos não funcionam de forma linear. von Békésy relata em seu O relato de que os potenciais intracelulares medidos nas células ci-
livro Experiments in Hearing (1960)39 que, quando a onda viajante liadas internas após um estímulo auditivo têm a mesma seletividade
foi observada pela primeira vez, ele não estava preocupado com a de frequências que as fibras do nervo coclear encerrou a discussão
amplitude dos movimentos. A intensidade do estímulo foi aumen- sobre o segundo filtro.44 Os resultados mostraram que uma análi-
tada até que a onda viajante fosse visível. A diminuição do estímulo se completa das frequências auditivas já existe nas células ciliadas
para metade da sua magnitude não alterava o padrão de vibração. internas e, portanto, “antes” do nervo auditivo.
Fig. 4-13. (a) Esquema ilustrando um instante (seta) da onda viajante que se propaga ao longo da membrana basilar em resposta a um tom puro. A cóclea,
que tem a forma de caracol, foi esticada e uma de suas divisões internas, a lâmina basilar, é mostrada como uma fita. Quando o amplificador coclear está
funcionando, as células ciliadas externas (CCE), numa região restrita próxima ao pico da onda, exercem uma força que amplifica o movimento da membrana
basilar no movimento que ocorreria, caso a mecânica coclear fosse passiva ou quando as células ciliadas externas não estão funcionando. (Modificada de
Ashmore.)40 (b) Esquema semelhante, porém observe que o pico da onda viajante é muito mais estreito do que o pico produzido durante a mecânica passiva,
estimulando uma área mais restrita da cóclea, o que é importante para a discriminação de frequências. O pico da onda viajante também ocorre numa região
mais basal do que o pico produzido pela mecânica passiva. (Modificada de Gummer e Preyer.)41
© 2021. Thieme. All rights reserved.
50 PARTE II SISTEMA AUDITIVO
Finalmente, metodologias mais sensíveis do que as empregadas CÓCLEA ATIVA
por Békésy, como a técnica de Mössbauer,45-47 cujo inventor interes- Em 1948, Gold já havia sugerido que, em virtude do alto grau de
santemente também ganhou o prêmio Nobel em 1961 (em Física), amortecimento existente no interior líquido da cóclea, a vibração
e a interferometria por laser48 foram capazes de demonstrar que os da partição coclear (lâmina basilar mais o órgão espiral) deveria
movimentos da lâmina basilar em um animal vivo diferem tanto requerer assistência mecânica dependente de energia metabólica,
quantitativa quanto qualitativamente dos movimentos anterior- a fim de manter a alta sensibilidade e a discriminação subjetiva de
mente observados post mortem. Para uma frequência específica, a frequências.49 Também imaginou que, se tal processo mecanicamen-
vibração no pico da onda viajante é muito mais localizada e apresen-
te ativo ocorresse, sons deveriam ser produzidos dentro da cóclea
ta uma amplitude consideravelmente maior. Além disso, a onda tem
como efeito colateral e poderiam ser captados no meato acústico
um comportamento de compressibilidade não linear, ou seja, com
externo. Essa foi a primeira ideia de que a orelha poderia emitir sons.
o aumento do estímulo, a resposta vibratória da membrana basilar
Gold tentou, inclusive, captar esses sons colocando microfo-
é cada vez menor, sugerindo que o processo funciona melhor com
nes próximo às orelhas, mas não obteve sucesso, principalmente
estímulos mais próximos ao limiar auditivo. Com sons de amplitude
pelas limitações tecnológicas da época. Suas conclusões, arrojadas
cada vez maior, a capacidade de discriminação de frequências se
demais para aquele momento, foram recebidas com descrédito e
deteriora, assemelhando-se aos resultados de von Békésy.
amplamente ignoradas. Seu trabalho ficou esquecido em meio a
Portanto, na cóclea viva, os movimentos da lâmina basilar são de
alguma forma amplificados, especialmente nos estímulos sonoros inúmeras outras “especulações” teóricas não experimentais sobre
próximos ao limiar auditivo, e são extremamente sintonizados às o funcionamento auditivo que apareceram nesse período. Gold che-
diversas frequências do estímulo. Esse ganho compensa o alto grau gou a visitar von Békésy, esperando que ele tivesse alguma “carta na
de amortecimento que existe no interior líquido da cóclea viva e é manga”, mas saiu convencido que von Békésy acreditava mesmo no
atualmente chamado de amplificador coclear, ou ainda de cóclea funcionamento passivo da cóclea e na inexistência de algum proces-
ativa. Por exemplo, no limiar auditivo, o movimento do pico ampli- so amplificador que compensasse o amortecimento da onda dentro
ficado da lâmina basilar é de aproximadamente 0,3 nm, chegando da cóclea. Esse desentendimento atrasou em aproximadamente
à excursão máxima de aproximadamente 50 nm. Sem esse ampli- 40 anos a descoberta das emissões otocústicas e, consequentemen-
ficador coclear, o movimento máximo seria aproximadamente 100 te, da triagem auditiva neonatal iniciada na década de 1990, que
vezes menor, ou seja, por volta de 0,003 nm (Fig. 4-13). tanto contribuiu e vem contribuindo para a identificação precoce
Hoje sabe-se que von Békésy descreveu o comportamento pas- das perdas auditivas neonatais.
sivo e linear da lâmina basilar, explicado pelas características físi- O papel das células ciliadas externas nesse processo começou a
cas dessa membrana e que ocorre na cóclea morta ou danificada. ser esclarecido quando vários grupos perceberam que a lesão des-
Entretanto, em um ser vivo, a alta capacidade de discriminação de sas células leva à deterioração da amplificação coclear. Assim, o
frequências não resulta apenas das propriedades vibratórias da lâ- que faz a diferença entre a cóclea ativa ou passiva é justamente a
mina basilar. presença das células ciliadas externas (Fig. 4-13). Atualmente, o
Dos experimentos de von Békésy, dois conceitos permanecem conhecimento do funcionamento da cóclea é caracterizado pelo
até hoje: a tonotopicidade e a onda viajante. Em ambos está a ideia conceito do amplificador coclear ou cóclea ativa, no qual a onda
de local equivalente à frequência, uma vez que uma onda viajante viajante de von Békésy é localmente amplificada por um processo
viaja em uma direção específica em razão da organização de fre- eletromecânico, em que as células ciliadas externas têm um papel
quências da membrana basilar. fundamental, exercendo as funções tanto de sensores como de ele-
Curiosamente, o próprio von Békésy entendeu melhor o con- mentos mecânicos de retroalimentação.
ceito de onda viajante do que muitos que seguiram suas ideias. Esses conceitos se fundamentaram na descoberta de David
Dois experimentos mostram fatos extremamente importantes so- Kemp de que a orelha interna emite sons espontaneamente e na
bre a onda viajante que costumam ser negligenciados em descri- observação de que as células ciliadas externas apresentam motili-
ções posteriores. dade, quando eletricamente estimuladas in vitro.50
O fato de a onda viajar da base ao ápice pode ter sido um re-
sultado inesperado, porém soa lógico. A onda inicia na base da có- EMISSÕES OTOACÚSTICAS
clea, pois é onde ocorre a introdução da energia desencadeada pelo
A descoberta de que sons provenientes da cóclea podiam ser re-
movimento da platina do estribo, e daí deve se distribuir ao longo
almente detectados no canal auditivo externo, tanto espontane-
da cóclea. No entanto, essa ideia não parece estar correta. Quando
amente como em resposta a um estímulo sonoro, como previsto
von Békésy estimulou as cócleas a partir da extremidade apical, a
por Gold, confirmou a presença de uma atividade motora dentro
onda viajante não foi afetada – ela continuou com o mesmo padrão,
da cóclea, pois tudo que se move, e não está no vácuo, produz sons.
viajando da base em direção ao ápice da cóclea!
Em outro experimento, ele fez um corte, uma interrupção, na Esse fenômeno, atualmente referido como emissões otoacústicas,
membrana basilar para examinar o grau em que a energia da onda é uma das mais interessantes descobertas que ocorreram nas últi-
é realmente transmitida ao longo das estruturas do órgão de Corti. mas décadas e revolucionou a compreensão do funcionamento do
Surpreendentemente, esses cortes não impediram a onda de viajar. sistema auditivo periférico.
Esses dois experimentos mostram, portanto, que: Essa descoberta foi recebida inicialmente com ceticismo, pois
se postulou que esses sons pudessem originar-se de estruturas da
1. A onda viaja da base ao ápice porque, após o estímulo pressórico orelha média. As primeiras evidências da origem intracoclear das
no estribo, as regiões mais apicais atrasam seu funcionamento emissões otoacústicas surgiram de experimentos mostrando que
em relação às regiões basais, ou seja, têm latências na resposta agentes que danificam especificamente as células ciliadas externas
ao estímulo sucessivamente maiores em relação à base; reduzem ou eliminam as emissões otocústicas. Exposição a ruído
2. Cada região do órgão espiral de Corti responde independente- e ototóxicos, como a furosemida e os aminoglicosídeos, são alguns
mente de seus vizinhos e com sua própria latência característica. exemplos de agentes que alteram as emissões otoacústicas. Por ou-
Talvez a onda viajante seja um epifenômeno, e não a causa do tro lado, o antineoplásico carboplatina, que, em chinchilas, induz
estímulo das células ciliadas. Pode-se observar uma onda “se mo- uma perda específica de células ciliadas internas, não causa dimi-
vendo”, mas a energia é fornecida por uma onda de pressão muito nuição nas emissões otoacústicas.
mais rápida que estimula global e diretamente a membrana basilar A demonstração definitiva da origem intracoclear das emissões
e o órgão de Corti. Se a onda viajante é decorrente das propriedades otoacústicas veio com os experimentos nos quais as emissões pu-
físicas da membrana basilar, como explicar a onda reversa que ocor- deram ser modificadas ou mesmo evocadas por meio de estímulo
re após a passagem pelo helicotrema em direção à janela redonda? elétrico no interior da cóclea ou sob estímulo eferente contralateral.
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 51
ELETROMOTILIDADE DAS CÉLULAS CILIADAS EXTERNAS esse fenômeno seja uma contração do tipo muscular, dependente
As células ciliadas externas foram apontadas como os efetores do de cálcio e de energia metabólica, envolvendo a interação de mo-
mecanismo ativo coclear após a demonstração de que apresentam léculas de actina e miosina.
alterações do seu comprimento em frequências correspondentes Entretanto, a eletromotilidade das células ciliadas externas é
às frequências auditivas, quando eletricamente estimuladas in vi- independente de cálcio e ATP,51 e a destruição do citoesqueleto e das
tro. Esses estudos demonstraram que, quando isolada em meio de cisternas endoplasmáticas não abole a motilidade, mostrando que a
motilidade das células ciliadas externas é um tipo de motilidade dis-
cultura, uma célula ciliada externa é despolarizada, ela se encurta
tinto da contração muscular. A atividade motora das células ciliadas
e se alarga. Quando é hiperpolarizada, se alonga e fica mais fina.
externas coincide com um denso arranjo de proteínas que se dis-
Essas alterações de comprimento podem chegar a 5% do com-
tribui ao longo da membrana plasmática basolateral (Fig. 4-14).52-55
primento da célula, o que representa uma enorme alteração em
A forma mais aceita de ação do motor é que essas proteínas
termos biológicos.
integrais de membrana, extensamente presentes na parede lateral
Várias técnicas biofísicas têm mostrado que o mecanismo ele-
das células ciliadas externas, e não nas células ciliadas internas, se-
tromotor pode ser acionado por toda a faixa de frequências que jam sensíveis às variações da voltagem da membrana plasmática.
abrangem a maioria dos órgãos auditivos de mamíferos e, pelo me- A alta densidade de prestina na membrana lateral da célula ci-
nos, até 80 kHz. liada externa é surpreendente. O empacotamento dessas proteínas
Acredita-se que o movimento conjugado de várias células cilia- na membrana plasmática da célula é apertado de tal forma que a
das externas acrescente energia mecânica aos movimentos da lâmi- densidade de partículas cobre cerca de 60% da superfície da parede
na basilar desencadeados pelo som, contribuindo para o estímulo lateral (Fig. 4-14). Essa aglomeração molecular amplifica qualquer
das células ciliadas internas. Como visto anteriormente, o proces- pequena alteração que ocorra na área de superfície de cada proteína
so ativo da cóclea é responsável pela sensibilidade auditiva e pela individual. Mesmo assim, para atuar como elemento gerador de for-
alta capacidade de discriminação de frequências apresentada pela ça, é necessário garantir que a membrana não se dobre, permitindo
orelha. A perda total da função das células ciliadas externas leva a que toda a força resultante seja gerada ao longo do eixo vertical da
uma perda auditiva de 50 a 60 dB. célula ciliada externa. Para garantir que a membrana mantenha a
Dados prévios demonstrando a presença de proteínas contrá- rigidez e a forma cilíndrica característica, permitindo a distribuição
teis, de um complexo citoesqueleto e de um arranjo de cisternas de força no eixo vertical, as células ciliadas externas desenvolveram
endoplasmáticas dentro dessas células, induziram à ideia de que uma rede cortical submembranosa.51
Fig. 4-14. O motor da célula ciliada externa. (a) Diagrama da membrana lateral da célula ciliada externa, onde os folhetos interno e externo da membrana
plasmática foram separados para revelar a alta densidade de partículas de membrana não presentes nas células ciliadas internas, que representam oligômeros
da proteína motora prestina. A rede cortical é composta de filamentos de actina paralelos orientados circunferencialmente ao redor da célula e reticulados
por espectrina. Os filamentos de actina estão ligados à membrana plasmática por moléculas pilares, e as cisternas laterais ficam logo dentro da rede cortical.
(Modificada de McGee e Walsh.)52 (b) Diagrama de uma pequena porção da parede lateral da célula ciliada externa com uma placa de oligômeros de prestina, as
quais sofrem mudanças conformacionais que alteram a densidade de empacotamento das partículas, em resposta a uma alteração na voltagem da membrana.
Durante a despolarização, um empacotamento mais apertado resulta no encurtamento dessa placa e, consequentemente, da própria célula ciliada; durante a
hiperpolarização, ocorre o alongamento. (Modificada de Frolenkov et al.)53 (c) Eletromicrografia de transmissão de réplica de carbono e platina por freeze-etching,
mostrando a estrutura da rede cortical. (Modificada de Holley et al.)54 (d) Em maior aumento, filamentos circunferenciais, pilares (P) e conexões são claramente
definidos. (Modificada de Holley et al.)54 (Continua)
© 2021. Thieme. All rights reserved.
52 PARTE II SISTEMA AUDITIVO
Fig. 4-14. (Cont.) (e) Eletromicrografia de transmissão por criofratura mostrando a alta densidade de proteínas integrais de membrana na face lateral da
membrana plasmática lateral (MPL) de célula ciliada externa. (Modificada de Kalinec et al.)55 (f) Réplicas por freeze-etching da membrana plasmática lateral
de uma célula ciliada externa mostrando a superfície mais externa da membrana, com um denso empacotamento de partículas, que posteriormente foram
identificadas como prestina. (Modificada de Kalinec et al.)55
O motor das células ciliadas externas é capaz de respostas ex-
tremamente rápidas, mais que qualquer outro motor biológico co-
nhecido, e não funciona somente quando se muda artificialmen-
te a voltagem por intermédio da membrana celular, mas também
quando se estimula mecanicamente o feixe estereociliar com um
jato de água ou com sons.
Um engenhoso trabalho de biologia molecular conseguiu iso-
lar o gene da proteína motora das células ciliadas externas, que foi
denominado prestina, em alusão ao movimento Presto da música
clássica.56 A estratégia utilizada foi a subtração de genes expressos
nas células ciliadas externas dos genes expressos nas células ciliadas
internas, uma vez que a célula ciliada interna não tem motilidade Fig. 4-15. Ilustração de moléculas de prestina localizadas na membrana
e não deve ter esse gene funcionando. Dessa subtração restaram plasmática que podem alternar entre dois estados conformacionais (alongado
ou encurtado), dependendo da voltagem da membrana plasmática; quanto
11 candidatos, sendo a prestina eleita como a provável proteína
maior a despolarização, maior a probabilidade de que o estado mais encolhido
motora das células ciliadas externas. Quando o gene da prestina foi seja adotado e vice-versa. Como a prestina é originária de uma família de
introduzido por meio de um vetor viral em culturas de células de proteínas de transporte iônico, acredita-se que ela seja um transportador
rim humano, essas células passaram a exibir as mesmas alterações incompleto. No modelo de controle de prestina por íons cloro intracelulares,
de comprimento induzidas por variações na voltagem da membrana os dois estados conformacionais seriam controlados pela ligação de íons
plasmática, característica das células ciliadas externas. cloro intracelulares. Na ausência de cloro, a molécula está em seu estado
Surpreendentemente, a prestina é membro de uma superfamí- “encolhido”. Quando a membrana celular é despolarizada, o cloro liga-se à
molécula, mas permanece na face citoplasmática da membrana. Quando a
lia de transportadores de membrana SLC26A5, uma família cujos
membrana celular é hiperpolarizada, o cloro migra através da molécula em
outros membros são trocadores de cloreto-bicarbonato.57 direção à face externa. Quando o cloro ligado à molécula está na posição mais
A questão de como uma proteína de transporte pode dar origem externa, a molécula assume sua conformação “alongada”. Quando o cloro é
a um motor foi resolvida com a proposta de que a prestina/SLC26A5 deslocado de volta para o citoplasma, a molécula volta ao estado “encurtado”.
seja um transportador incompleto. A ideia é que a proteína atue como (Modificada de Dallos e Fakler.)57
uma mecanoenzima como parte de seu ciclo de transporte, porém
sem fazer transporte. Com a ligação de um ânion intracelular (clo-
para a rigidez das células ciliadas externas, e que a remoção da pres-
reto), ocorre uma mudança conformacional na proteína, suficiente
tina altera o acoplamento do som à mecânica da membrana basilar.
para produzir alteração de comprimento no plano da membrana
e, portanto, no comprimento da célula ciliada externa (Fig. 4-15).58 Essa questão foi contornada usando-se um camundongo com
Tem havido alguma controvérsia sobre a prevalência de mu- mutações não desestruturantes da prestina, em que as células ci-
tações prestina/SLC26A5 em humanos. Até agora as mutações da liadas externas são estrutural e biofisicamente próximas do nor-
prestina humana parecem bastante raras. Um único relato de caso mal, mas têm propriedades elétricas alteradas.62 Nesse mutante, de
identifica duas irmãs profundamente surdas com mutações hete- fato, a membrana basilar não está sintonizada e há uma redução na
rozigotas compostas p.W70X e p.R130S.59 seletividade de frequência e na sensibilidade coclear, mesmo sem
Em camundongos, há ampla evidência de que a prestina real- esse desacoplamento da mecânica coclear.
mente determine a sensibilidade coclear. Os primeiros knockouts de
prestina mostraram que as células ciliadas externas isoladas desses Motilidade dos Estereocílios
camundongos não são móveis e que há uma perda de sensibilidade
Martin e Hudspeth propuseram a existência de um processo motor
coclear de 40-60 dB in vivo.60 O microfonismo coclear ainda podia ser
localizado nos canais de transdução que serviria para amplificar
medido, mostrando que a transdução mecanoelétrica não é afetada,
a transdução da célula ciliada.63 Essa motilidade ativa nos estere-
embora pareça haver apoptose progressiva das células.
Uma observação direta do movimento da membrana basilar na ocílios ainda não foi observada nas células ciliadas externas, mas
região mais basal da cóclea, correspondente às frequências de 60 a possivelmente pode coexistir com a motilidade somática observada
70 kHz, também provou que a alta capacidade de sintonia mecânica nos mamíferos. Acredita-se que a entrada de cálcio pelos canais de
de camundongos normais está ausente no camundongo knockout de transdução desencadeie a movimentação ativa dos estereocílios.
prestina. Contudo, curiosamente, o limiar auditivo não é significati- Esse amplificador estereociliar explicaria a presença de emis-
vamente rebaixado.61 A possível explicação é que a prestina contribua sões otoacústicas em animais sem células ciliadas externas, como
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 53
anfíbios, aves e répteis, e cujas células ciliadas não apresentam ciclos Outro potencial de repouso é o intracelular, que é o potencial
de alongamento e encurtamento (motilidade somática). negativo existente no interior das células do órgão espiral. O po-
tencial das células ciliadas externas é, em média, -70 mV, próximo
Potenciais Cocleares ao das células de suporte, que varia de -70 a -100 mV. Já as células
As interações entre as várias estruturas cocleares durante o processo ciliadas internas possuem potencial na ordem de -45 mV.
auditivo geram potenciais elétricos que podem ser captados e estu- Como mencionado anteriormente, a diferença entre o potencial
dados. Não se sabe ao certo se esses potenciais são sempre respostas intracelular de -70 mV nas células ciliadas e o potencial de +80 mV
fisiológicas ou simplesmente efeitos colaterais da atividade coclear. da endolinfa gera uma diferença de 150 mV, que representa a maior
Existem duas classes de potenciais: os potenciais que ocorrem diferença de potencial encontrada em toda a natureza.
após um estímulo acústico, ou dependentes do estímulo, e os de Além do potencial endolinfático e do potencial de repouso intra-
celular, há ainda um terceiro potencial de repouso, de importância
repouso, que independem do estímulo. Os potenciais cocleares po-
e origem desconhecidas. É uma pequena diferença de potencial de
dem ser medidos nas células do labirinto membranoso e nos es-
2 a 5 mV entre a rampa do vestíbulo e a rampa do tímpano.
paços cocleares.
Até recentemente, o entendimento do funcionamento cocle-
ar baseou-se, principalmente, no estudo da atividade elétrica dos
Potenciais Dependentes do Estímulo Acústico
compartimentos intra e extracelulares do labirinto membranoso. As respostas elétricas desencadeadas pelo estímulo acústico são
geradas pelas células ciliadas do órgão espiral. Esses potenciais são
Atualmente, diversas outras técnicas de fisiologia quase suplanta-
medidos por meio de eletrodos posicionados na rampa média e re-
ram os estudos dos potenciais cocleares, embora a eletrococleografia
presentam a média das respostas de várias células ciliadas ao longo
ainda seja utilizada na prática clínica.
da lâmina basilar.
Os potenciais evocados por um tom puro possuem dois com-
Potenciais de Repouso ponentes: uma corrente fásica ou alternada, cuja frequência é a
Existem dois potenciais de repouso de maior importância. O po- mesma do estímulo, e um componente de corrente contínua, que
tencial endococlear ou endolinfático, que é medido no interior da desloca o potencial de base, o potencial endolinfático, em direção
rampa média, sendo positivo em relação à perilinfa das rampas do negativa, durante o estímulo. A resposta fásica é o microfonismo
tímpano e do vestíbulo. É um potencial de corrente contínua (cons- coclear e a resposta contínua, o potencial de somação. O micro-
tante) de aproximadamente +80 mV na base e ligeiramente menor fonismo coclear representa a corrente elétrica resultante da en-
em direção à cúpula da cóclea. trada de potássio nas células ciliadas pelos canais de transdução.
Acredita-se que o potencial endolinfático seja produzido pela A exata origem do potencial de somação não é conhecida, mas hoje
estria vascular, pelo transporte ativo de potássio para o interior do se acredita que seja o potencial que ocorre na despolarização das
ducto coclear em troca de sódio. Essa troca seria realizada pelas cé- células ciliadas internas.
lulas marginais, ricas em sódio-potássio adenosina trifosfatase (Na+/
K+ ATPase), mantendo, dessa forma, a concentração de potássio no Fisiologia do Sistema Eferente Coclear
interior da rampa média em torno de 144 mEq/L em comparação com Uma rica população de fibras nervosas eferentes, paralela às vias
4 mEq/L da rampa do vestíbulo. O potencial endolinfático é o potencial ascendentes, comunica o córtex auditivo com os centros auditivos
mais positivo encontrado em todo o corpo humano. Não é encontrado inferiores e com o órgão espiral.
na endolinfa do sistema vestibular, apesar do fluxo de endolinfa ser Este sistema eferente, descrito inicialmente por Rasmussen, é
comunicante em todo o labirinto. Essa seria uma das evidências de composto pelo sistema olivococlear medial e o sistema olivococle-
que o potencial endolinfático é gerado pela estria vascular, que está ar lateral (Fig 4-16a).64 O sistema olivococlear medial origina-se no
presente somente nas paredes laterais do ducto coclear. subnúcleo do complexo olivar medial superior, projeta-se através de
Fig. 4-16. Esquema mostrando a inervação aferente (em azul) e eferente (em vermelho) do órgão espiral de Corti. (a) Distribuição das fibras aferentes do tipo
I e do tipo II dos neurônios do gânglio espiral e os sistemas eferentes medial e lateral. Enquanto 10 a 20 fibras aferentes do tipo I fazem conexões diretas com
apenas 1 célula ciliada interna, uma única fibra aferente do tipo II ramifica-se e faz contato com várias células ciliadas externas. Essas ramificações são sempre
em direção à base da cóclea. (b) As linhas vermelhas conectando neurônios do núcleo olivococlear lateral representam fibras finas não mielinizadas; linhas azuis
conectando neurônios olivococlear medial representam fibras mielinizadas. O sistema eferente faz terminações diretas com os corpos celulares das células
ciliadas externas e faz apenas terminações en passant com as fibras aferentes, abaixo das células ciliadas internas, no feixe espiral interno. As fibras originárias
do núcleo olivococlear lateral projetam-se predominantemente ipsilateralmente enquanto as fibras originárias de neurônios do núcleo olivococlear medial
projetam-se predominantemente para o lado contralateral. Os aferentes cruzam o túnel de Corti no assoalho enquanto os eferentes cruzam na porção média do
túnel. OC: olivococlear; CCE: células ciliadas externas; CCI: célula ciliada interna. (Modificada de Schuknecht.)64
© 2021. Thieme. All rights reserved.
54 PARTE II SISTEMA AUDITIVO
fibras mielinizadas predominantemente para a cóclea contralateral o feedback eferente para a cóclea seja plástico, tornando-se mais
e termina diretamente nas células ciliadas externas (Fig. 4-16b). O forte com o uso e treinamento.
sistema olivococlear lateral origina-se em pequenos neurônios no Tem sido difícil associar o mau funcionamento de neurônios
núcleo olivar superior lateral e é formado por fibras não mieliniza- olivococleares a doenças. Mesmo assim há algumas evidências de
das que se projetam predominantemente para a cóclea ipsilateral atividade olivococlear anormal em pessoas com zumbido, hipera-
e terminam quase que exclusivamente nos terminais nervosos das cusia e distúrbios do processamento auditivo central.70 O proces-
células ciliadas internas. Não fazem contato direto com as células samento eferente anormal foi também postulado em uma série de
ciliadas internas.64,65 distúrbios neurológicos que cursam com processamento auditivo
Os sistemas eferentes, medial e lateral têm padrões distintos desregulado: autismo, esquizofrenia, enxaqueca, mutismo seletivo,
de inervação ao longo da espiral coclear. A inervação eferente la- comprometimento específico da linguagem e dislexia.71,72
teral projeta-se de forma uniforme por toda a cóclea, enquanto a
inervação eferente medial é maior na base da cóclea, decrescendo Sistema Aferente
em direção à cúpula e maior na primeira fileira de células cilia- Enquanto a frequência do som é codificada, principalmente, pela
das externas. posição de uma célula ciliada ao longo do comprimento da cóclea,
Embora os resultados sobre a significância fisiológica do sistema a intensidade e a análise temporal da onda sonora são definidas
eferente sejam conflitantes, várias ações desse sistema têm sido re- pelo padrão de disparo do nervo auditivo, controlado pela libera-
latadas e indicam um efeito inibitório sobre as respostas cocleares. ção precisa de neurotransmissores nas sinapses das células ciliadas
Seu efeito mais conhecido é obtido por meio da estimulação internas.73
elétrica do feixe olivococlear no assoalho do quarto ventrículo, re- Os dois tipos de células ciliadas têm papéis complementares
sultando em diminuição da amplitude e aumento na latência do po- na transdução sensorial auditiva, mas sua inervação é totalmen-
tencial de ação composto do nervo auditivo. Outros efeitos relatados te distinta. A fileira única de células ciliadas internas retransmite
são o aumento do microfonismo coclear, a alteração nas amplitudes todas as informações sonoras para fibras aferentes tipo I do nervo
e nas frequências dos produtos de distorção das emissões otoacús- auditivo por meio da liberação de vesículas sinápticas contendo
ticas, além da diminuição na sensibilidade e na discriminação de glutamato. As fibras do tipo I constituem a maioria dos neurônios
frequências nas curvas de sintonia do nervo auditivo. do gânglio espiral (NGE) que inervam a cóclea (aproximadamente,
As vias descendentes do feixe olivococlear medial liberam ace- 95%). Por outro lado, as três fileiras de células ciliadas externas au-
tilcolina (ACh) e ativam receptores de acetilcolina heteroméricos mentam a sensibilidade e as propriedades de ajuste de frequência
α9/10 (diferente de todos os receptores de acetilcolina no organis- da partição coclear.
mo) nas células ciliadas externas. Servem para controlar a condutân- Embora as células ciliadas externas sejam principalmente iner-
cia da membrana da célula para o potássio, através da ativação de vadas por neurônios colinérgicos eferentes, estas recebem também
canais de potássio cálcio-dependentes. O efeito final é hiperpolari- inervação de neurônios aferentes do tipo II que compõem os ∼5%
zar as células ciliadas externas e reduzir a amplitude dos potenciais restantes dos neurônios aferentes do gânglio espiral.
da recepção. Consequentemente, a eletromotilidade da célula ciliada Em contraste com as fibras do tipo I, que fazem contato sináp-
externa é suprimida, reduzindo a amplificação do movimento da tico com apenas uma célula ciliada interna, os terminais periféricos
membrana basilar e o ganho mecânico coclear geral. dos neurônios aferentes do tipo II formam extensas arborizações em
A observação de que as células ciliadas externas da primeira espiral com várias células ciliadas externas (Fig. 4-16a). A função
fileira recebem consideravelmente mais contatos eferentes do que das fibras do tipo II ainda é desconhecida, embora estas pareçam
as células das duas outras fileiras sugere que a primeira coluna deve ser ativadas apenas por sons muito altos, o que levou à suposição
contribuir mais para as pontas das curvas de sintonia (discriminação de que funcionem como nociceptores cocleares, alertando e evi-
fina de frequências) do que a segunda e terceira fileiras. tando a piora da lesão.74
Os neurônios do sistema lateral olivococlear também modu-
lam os aferentes do tipo I que saem das células ciliadas internas Sinapse da Célula Ciliada Interna
por mecanismos sinápticos ainda desconhecidos. Os efeitos dessa Desde a virada deste século, tanto o uso de imagens de alta resolução
modulação também ainda não são entendidos. Tem sido mais difícil das sinapses das células ciliadas internas, como o desenvolvimento
determinar os efeitos dos eferentes do sistema lateral. Experimentos de novas técnicas eletrofisiológicas forneceram muitas informações
in vivo produziram resultados mistos, possivelmente refletindo a sobre essa sinapse (Fig. 4-17). Em paralelo, o estudo das mutações
neuroquímica mais complexa desse sistema, que inclui dopamina genéticas alavancou o conhecimento da arquitetura molecular e da
e GABA, além da própria acetilcolina.66 função dessa sinapse.
O papel do sistema eferente é controverso, mas seu efeito so- A abertura dos canais de transdução mecanoelétrica no polo
bre o ajuste mecânico67 e as emissões otoacústicas de produtos de apical das células ciliadas causa a conversão (transdução) da infor-
distorção68 é claro. Há também a sugestão, a partir de estudos em mação acústica em um potencial de recepção. Esse potencial ativa
camundongos nos quais o receptor colinérgico eferente foi excluído, canais de cálcio do tipo L, dependentes de voltagem (CaV1.3), locali-
de que a atividade do sistema eferente pode até retardar o envelhe- zados nas zonas ativas pré-sinápticas no polo basal da célula ciliada
cimento auditivo.69 Esse feedback negativo também pode proteger (Fig. 4-17). A entrada de cálcio resultante desencadeia a fusão de
a cóclea contra trauma acústico. Vários trabalhos sugerem ainda vesículas sinápticas, liberando glutamato nos botões pós-sinápticos
que os eferentes podem ser ativados pelo sistema nervoso central, dos neurônios do gânglio espiral.75-77
podendo exercer um papel na atenção seletiva, melhorar a codifi- No citoplasma adjacente à zona de liberação, na base das cé-
cação do sinal em ambientes com muito ruído, modular a atenção lulas ciliadas internas, um grande número de vesículas sinápticas
seletiva aos estímulos sensoriais e influenciar a localização sonora. contendo glutamato aparecem ligadas a estruturas elétron-densas
Os eferentes olivococleares desempenham também um papel chamadas corpos pré-sinápticos, de formato esférico ou ovoide,
ainda não totalmente compreendido na maturação da inervação com cerca de 400 nm de diâmetro (Fig. 4-17),78 e que servem como
coclear. As sinapses inibitórias do sistema lateral fazem contato di- reservatório de neurotransmissores.
reto com as células ciliadas internas apenas no início embrionário A liberação de neurotransmissor da sinapse da célula ciliada in-
da audição, regulando a atividade espontânea de neurônios aferen- terna é desencadeada por variações graduais do potencial de mem-
tes, o que molda a conectividade dos núcleos do tronco cerebral. brana, aumentando com a intensidade do som. A resposta sináp-
O papel fisiológico do feedback eferente na função auditiva tem tica aos sons de até 3 kHz ocorre em fase com a onda de pressão
se mostrado difícil de compreender. As medições da sua função sonora. Isso implica numa alta precisão temporal de transmissão
mostram, às vezes, efeitos contrários ou nenhum efeito, em estu- sináptica, essencial para a percepção do pitch e da localização da
dos diferentes. Isso pode refletir, em parte, a possibilidade de que fonte sonora.79
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 55
Fig. 4-17. Configuração das sinapses entre a célula ciliada interna e a fibra aferente coclear. (a) Cada sinapse tem um corpo pré-sináptico (ribbon) próximo à
área de liberação de neurotransmissores e adjacente à membrana pós-sináptica do neurônio aferente. No lado modiolar, as regiões pré-sinápticas têm corpos
pré-sinápticos maiores (vermelho) e exibem entradas de cálcio maiores (azul claro). Os neurônios aferentes têm diâmetros menores, limiar de sensibilidade
maior e taxa de disparos espontâneos menores. No lado do pilar, os ribbons são menores e o influxo de cálcio é menor; os aferentes têm diâmetro maior, limiar
mais baixo e maior taxa de disparos espontâneos. (Modificada de Johnson et al.)74 (b) Eletromicrografia de transmissão mostrando o corpo pré-sináptico em
grande aumento, circundado de vesículas sinápticas. (Modificada de Safieddine et al.)75 (c, d) O corpo pré-sináptico é composto pelas proteínas Ribeye e Piccolo
e ancorada à membrana do local de liberação pela proteína Bassoon. As vesículas circundantes (em amarelo) contêm o neurotransmissor glutamato e são
exocitadas pelo influxo de cálcio através dos canais de cálcio Cav1.3 na membrana pré-sináptica. O glutamato se liga aos receptores ionotrópicos AMPA GluA2/3
na membrana pós-sináptica. Visão em maior detalhe da vesícula sináptica, mostrando o transportador de glutamato Vglut3 e o sensor de cálcio otoferlina.
(Modificada de Fettiplace.)76
Parte dos canais de transdução mecanoelétrica estão sempre loss), um distúrbio auditivo recentemente descrito em que algumas
abertos, mesmo sem estímulo sonoro, resultando na liberação es- pessoas têm limiares auditivos normais, mas dificuldade de perce-
pontânea de glutamato. Essa liberação de neurotransmissores em ção significativa, principalmente em ambientes ruidosos.
repouso nas células ciliadas internas modula a taxa de potenciais Diferentemente das sinapses convencionais do SNC, as sinapses
de ação espontâneos (APs) nos NGE aferentes tipo I. das células ciliadas internas expressam otoferlina (Fig. 4-17c), que
Cada célula ciliada interna é inervada por 10 a 30 neurônios funciona como o principal sensor de Ca+2 para a exocitose,80 além
aferentes bipolares do tipo I não ramificados e que formam peque- de estar envolvida em outras funções do ciclo da vesícula sináptica.
nos contatos ou botões adjacentes a um único corpo pré-sináptico O cálcio nas zonas pré-sinápticas liga-se à otoferlina e dispara
(Figs. 4-16 e 4-17). Em camundongos, o número de sinapses nas a fusão das vesículas sinápticas contendo glutamato com a mem-
células ciliadas internas varia tonopicamente, com menos contatos brana plasmática pré-sináptica. A otoferlina, como sensor de Ca+2,
no ápice e na base coclear e mais sinapses na região central da có- determina a taxa e a concentração necessárias de Ca+2 para desen-
clea, que corresponde à região mais sensível e mais utilizada para cadear a fusão e a reposição das vesículas sinápticas.
a comunicação e vocalização, como se fosse uma fóvea auditiva. A otoferlina foi identificada como defeituosa na DFNB9, uma
As terminações aferentes das células ciliadas internas apresen- forma recessiva de surdez pré-lingual humana.81 A similaridade
tam diferentes características morfofuncionais. As sinapses no lado nos resultados dos testes audiológicos entre pacientes DFNB9 e
modiolar das células ciliadas internas geralmente têm neurônios camundongos deficientes em otoferlina (OTOF -/-), juntamente
aferentes com diâmetros menores, sinapses com corpos pré-sináp- com a análise fisiopatológica realizada nesses camundongos OTOF
ticos maiores, botões pós-sinápticos de receptores AMPA menores, -/-, sugere que a surdez DFNB9 é causada por um defeito primário
limiar mais alto de sensibilidade, taxa de disparo espontânea mais na fusão vesicular na sinapse da célula ciliada interna. Esse e ou-
baixa e entradas de cálcio maiores (Fig. 4-17a). tros estudos posteriores revelaram a sinaptopatia como um dos
Do lado do pilar, as sinapses exibem características opostas, principais mecanismos que levam à surdez humana DFNB9.80 Pelo
ou seja, neurônios com diâmetros maiores, corpos pré-sinápticos que se sabe até hoje, neuropatia auditiva é a única manifestação
menores com menor entrada de cálcio, botões pós-sinápticos maio- fenotípica de mutações no gene OTOF.
res, limiar de sensibilidade mais baixo, taxa de disparo espontânea O termo neuropatia auditiva foi cunhado pela primeira vez
mais alta (Fig. 4-17a). por Starr et al.82 Atualmente denominada de Transtorno do Espec-
As sinapses do lado modiolar são as primeiras a ser lesadas tro da Neuropatia Auditiva, é um tipo de perda auditiva em que os
na exposição temporária ao ruido e no envelhecimento, podendo limiares auditivos tonais são normais ou quase normais e os refle-
explicar alguns dos casos de perda auditiva oculta (hidden hearing xos acústicos e os potenciais evocados do tronco encefálico (PEA-
© 2021. Thieme. All rights reserved.
56 PARTE II SISTEMA AUDITIVO
TE) estão ausentes ou muito alterados. A função das células ciliadas 13. Beurg M, et al. Localization of inner hair cell mechanotransducer
externas é normal, conforme indicado pela presença de emissões channels using high-speed calcium imaging. Nat Neurosc.
otoacústicas e/ou microfonismo coclear. Essa combinação de re- 2009;12(5):553-558.
14. Corey DP, Hudspeth AJ. Response latency of vertebrate hair cells.
sultados indica que a via auditiva até, e incluindo, a célula ciliada
1979;26(3):499-506.
externa está funcionando, mas o estímulo auditivo não é transmi- 15. Holt JR, Corey DP. Two mechanisms for transducer adaptation in
tido ao tronco cerebral, sugerindo que a lesão pode ser nas células vertebrate hair cells, (X). 2000.
ciliadas internas, na sinapse das células ciliadas internas para as 16. Kachar B, et al. High-resolution structure of hair-cell tip links.
fibras aferentes ou no próprio nervo auditivo. Indivíduos com esse Proceedings of the National Academy of Sciences of the United States
transtorno podem ter vários graus de perda auditiva, conforme me- of America. 2000;97(24):13336-13341.
dido pela audiometria tonal. Eles geralmente têm compreensão de 17. Kazmierczak P, et al. Cadherin 23 and protocadherin 15
fala desproporcionalmente pobre. A maioria dos indivíduos não se interact to form tip-link filaments in sensory hair cells. Nature.
2007;449(7158):87-91.
beneficia do uso de aparelhos auditivos, mas pode ser ajudada por
18. Sotomayor M, et al. Structure of a force-conveying cadherin bond
implantes cocleares, principalmente quando a causa da neuropatia essential for inner-ear mechanotransduction. Nature. 2012:128-132.
é a sinapse da célula ciliada interna. 19. Furness DN, Hackney CM. Cross-links between stereocilia in the
guinea pig cochlea. Hearing Res. 1985;18(2):177-188.
CONSIDERAÇÕES FINAIS 20. Jacobs RA, Hudspeth A J. Ultrastructural correlates of
A anatomia microscópica da orelha interna, a importância das célu- mechanoelectrical transduction in hair cells of the bullfrog’s internal
ear, in Cold Spring Harbor Symposia on Quantitative Biology. 1990.
las ciliadas e, especialmente, o reconhecimento das células ciliadas
21. Fettiplace R. Is TMC1 the Hair Cell Mechanotransducer Channel?.
internas como verdadeiros receptores do estímulo auditivo são co-
Biophy J. 2016;111(1):3-9.
nhecidos há bastante tempo. 22. Gillespie PG, Müller U. Mechanotransduction by Hair Cells: Models,
A descoberta da motilidade das células ciliadas externas, o co- Molecules, and Mechanisms. Cell. 2009:33-44.
nhecimento da sua ultraestrutura e seu reconhecimento na poten- 23. Wu Z, et al. Mechanosensory hair cells express two molecularly
cialização do estímulo auditivo trouxeram uma verdadeira revolu- distinct mechanotransduction channels. Nat Publi Group.
ção no campo da otologia. As emissões otoacústicas confirmaram 2017;20(1):24-33.
a existência de um processo ativo, mediado pelas células ciliadas 24. Kawashima Y, et al. Mechanotransduction in mouse inner ear hair
cells requires transmembrane channel-like genes. J Clin Invest.
externas, dentro da cóclea e têm sido muito utilizadas clinicamente
2011;121(12):4796-4809.
na identificação precoce da perda auditiva neonatal.
25. Pan B, et al. TMC1 and TMC2 are components of the
Mais recentemente, a codificação do genoma humano e as téc- mechanotransduction channel in hair cells of the mammalian inner
nicas de sequenciamento genético de última geração têm propi- ear. Neuron. 2013;79(3):504-515.
ciado a identificação de genes que causam a perda auditiva e os 26. Beurg M, Goldring AC, Fettiplace R. The effects of Tmc1 Beethoven
respectivos produtos desses genes. Isso vem proporcionando um mutation on mechanotransducer channel function in cochlear hair
conhecimento quanto ao funcionamento do processo auditivo ao cells. J General Phys. 2015;146(3):233-243.
nível molecular nunca imaginado. 27. Kurima K, et al. TMC1 and TMC2 Localize at the Site of
Mechanotransduction in Mammalian Inner Ear Hair Cell Stereocilia.
Atualmente, para se descrever a fisiologia auditiva, é preciso
Cell Reports. Authors. 2015;12(10):1606-1617.
se aprofundar nas complexas interações químicas dos personagens
28. Corns LF, et al. Tmc1 point mutation affects Ca2+ sensitivity and block
moleculares que compõem o órgão auditivo. by dihydrostreptomycin of the mechanoelectrical transducer current
Por outro lado, esse conhecimento propiciará em breve a desco- of mouse outer hair cells. J Neurosc. 2016;36(2):336-349.
berta de tratamentos moleculares por terapia genética, ou mesmo 29. Jia Y, et al. TMC1 and TMC2 Proteins Are Pore-Forming Subunits of
por drogas, das perdas auditivas, que por muito tempo denomina- Mechanosensitive Ion Channels. Neuron. Cell Press. 2020;105(2):310-
mos de irreversíveis. 321.e3.
Estamos na vigência de uma grande e benvinda revolução no 30. Maeda R, et al. Tip-link protein protocadherin 15 interacts with
entendimento do funcionamento auditivo normal e consequen- transmembrane channel-like proteins TMC1 and TMC2 Proceedings
of the National Academy of Sciences of the United States of America.
temente no tratamento das doenças que afetam a orelha interna.
2014;111(35):12907-12912.
31. Nist-Lund CA, et al. Improved TMC1 gene therapy restores hearing
REFERÊNCIAS BIBLIOGRÁFICAS and balance in mice with genetic inner ear disorders. Nature
1. Merchant SN, et al. Analysis of middle ear mechanics and application Communications. Springer US. 2019;10(1):1-14.
to diseased and reconstructed ears. Am J Otol. 1997;18(2):139-154. 32. Li X, et al. Localization of TMC1 and LHFPL5 in auditory hair cells in
2. Fettiplace R, Kim KX. The physiology of mechanoelectrical neonatal and adult mice. FASEB J. 2019.
transduction channels in hearing. Physiological Reviews. Am Physiol 33. Asai Y, et al. Transgenic Tmc2 expression preserves inner ear hair
Society. 2014:951-986. cells and vestibular function in mice lacking Tmc1. Scientific Reports.
3. Corey DP, Hudspeth AJ. Response latency of vertebrate hair cells. 2018;8(1):1-15.
Biophys J. 1979;26(3):499-506. 34. Nakanishi H, et al. Tmc2 expression partially restores auditory
4. Hudspeth AJ. Integrating the active process of hair cells with function in a mouse model of DFNB7/B11 deafness caused by loss of
cochlear function. Nature Reviews Neuroscience. Nat Pub Group. Tmc1 function. Scientific Reports. 2018;8(1):1-13.
2014:600:614. 35. Dionne G, et al. Mechanotransduction by PCDH15 Relies on a Novel
5. Dallos P. Cochlear neurobiology: revolutionary developments. ASHA. cis-Dimeric Architecture. Neuron. Cell Press. 2018;99(3):480-492.e5.
1988;30(6-7):50-56. 36. Pan B, et al. TMC1 Forms the Pore of Mechanosensory Transduction
6. Fawcett D. The Cell. Philadelphia: WB Saunders; 1981. Channels in Vertebrate Inner Ear Hair Cells. Neuron. Cell Press.
7. Frolenkov GI, et al. Genetic insights into the morphogenesis of inner 2018;99(4):736-753.e6.
ear hair cells. Nature Reviews Genetics. 2004;5(7):489-498. 37. Lichtenwanger W, von Helmholtz H, Ellis AJ. On the Sensations of Tone
8. Richardson GP, Petit C. Hair-bundle links: Genetics as the gateway to as a Physiological Basis for the Theory of Music. Notes. 1954.
function, Cold Spring Harbor Perspectives in Medicine. Cold Spring 38. Kursell J. Alexander ellis’s translation of helmholtz’s sensations of
Harbor Laboratory Press. 2019;9(12). tone. Isis. 2018;109(2):339-345.
9. Assad JA, Shepherd GMG, Corey DP. Tip-link integrity and mechanical 39. von Békésy G. Experiments in Hearing, McGraw-Hill Book Company,
transduction in vertebrate hair cells. Neuron. 1991. Inc; 1960.
10. Basu A, et al. Direct mechanical stimulation of tip links in hair cells 40. Ashmore J F. Hair cell based amplification in the cochlea. Curr Opinion
through DNA tethers. eLife; 2016. Neurobiol. 1994;4(4):503-508.
11. Pickles JO, Comis SD, Osborne MP. Cross-links between stereocilia in 41. Gummer AW, Preyer S. Cochlear amplification and its pathology:
the guinea pig organ of Corti, and their possible relation to sensory Emphasis on the role of the tectorial membrane. Ear Nose Throat J.
transduction. Hearing Research; 1984. 1997;76(3):151-163.
12. Corey DP, Hudspeth AJ. Ionic basis of the receptor potential in a 42. Moore BCJ. Contributions of von Békésy to psychoacoustics”. Hear Res.
vertebrate hair cell. Nature. 1979;281:675-677. 2012;293:51-7.
© 2021. Thieme. All rights reserved.
CAPÍTULO 4 FISIOLOGIA DO SISTEMA AUDITIVO 57
43. Evans EF. The frequency response and other properties of single fibres of the National Academy of Sciences of the United States of America.
in the guinea-pig cochlear nerve. J Physiol. 1972;226(1):263-287. 1999;96(25):14306-14311.
44. Russell IJ, Sellick PM. Intracellular studies of hair cells in the 64. Rasmussen GL. The olivary peduncle and other fiber projections of the
mammalian cochlea. J Physiol. 1978. superior olivary complex. J Comp Neurol. 1946;84:141-219.
45. Johnstone B M, Boyle A J F. Basilar membrane vibration examined with 65. Schuknecht HF. Pathology of the ear. 2nd ed. Philadelphia: Lea &
the Mössbauer technique. Science. 1967;158(3799):389-390. Febiger. 1993;31-75.
46. Rhode WS. Observations of the Vibration of the Basilar Membrane in 66. Ruel J, et al. Physiology, pharmacology and plasticity at the inner hair
Squirrel Monkeys using the Mössbauer Technique. J Acoust Society cell synaptic complex. Hearing Research. 2007;227(1-2):19-27.
Am. 1971. 67. Murugasu E, Russell IJ. The effect of efferent stimulation on basilar
47. Sellick PM, Patuzzi R, Johnstone BM. Measurement of basilar membrane displacement in the basal turn of the guinea pig cochlea.
membrane motion in the guinea pig using the Mössbauer technique. J Neurosc. 1996;16(1):325-332.
J Acoust Society Am. 1982. 68. Maison SF, et al. Overexpression of SK2 channels enhances efferent
48. Khanna SM, Leonard DGB. Basilar membrane tuning in the cat suppression of cochlear responses without enhancing noise
cochlea. Science. 1982. resistance. J Neurophysiol. 2007;97(4):2930-2936.
49. Gold T. Hearing. II. The physical basis of the action of the 69. Charles Liberman M, Liberman L D, Maison S F. Efferent feedback
cochlea. Proceedings of the Royal Society B: Biological Sciences. slows cochlear aging. J Neurosc. 2014;34(13):4599-4607.
1948;135(881):492-498. 70. Knudson IM, Shera CA, Melcher JR. Increased contralateral
50. Kemp DT. Stimulated acoustic emissions from within the human suppression of otoacoustic emissions indicates a hyperresponsive
auditory system. J Acoust Soc Am. 1978;64(5):1386-1391. medial olivocochlear system in humans with tinnitus and
51. Kalinec F, et al. A membrane-based force generation mechanism in hyperacusis. J Neurophysiol. 2014;112(12):3197-3208.
71. Khalfa S, et al. “Peripheral auditory asymmetry in infantile autism”,
auditory sensory cells. Cell Biology. 1992.
European Journal of Neuroscience. 2001;13(3):628-632.
52. McGee J, Walsh EJ. Cochlear Transduction and the Molecular Basis of
72. Veuillet E, et al. Abnormal peripheral auditory asymmetry in
Auditory Pathology. Seventh Ed, Cummings Otolaryngology – Head
schizophrenia. J Neurol Neurosurg Psychiat. 2001;70(1):88-94.
and Neck Surgery. Seventh Ed. Elsevier Inc; 2010.
73. Johnson SL, et al. Hair cell afferent synapses: Function and
53. Frolenkov GI, et al. The membrane-based mechanism of cell motility
dysfunction. Cold Spring Harbor Perspectives in Medicine. Cold Spring
in cochlear outer hair cells. Mol Biol Cell. 1998/9(8):1961-1968.
Harbor Laboratory Press. 2019;9(12).
54. Holley MC, Kalinec F, Kachar B. Structure of the cortical cytoskeleton
74. Liu C, Glowatzki E, Fuchs PA. Unmyelinated type II afferent neurons
in mammalian outer hair cells. J Cell Science. 1992;102(3):569-580.
report cochlear damage. Proceedings of the National Academy of
55. Kalinec F, et al. A membrane-based force generation mechanism in
Sciences of the United States of America. 2015;112(47):14723-14727.
auditory sensory cells. Cell Biology; 1992.
75. Johnson SL, et al. Hair cell afferent synapses: Function and
56. Zheng J, Shen W, He DZ, et al. Prestin is the motor protein of cochlear dysfunction. Cold Spring Harbor Perspectives in Medicine. Cold Spring
outer hair cells. Nature. 2000;405(6783):149-155. Harbor Laboratory Press. 2019,9(12).
57. Lohi H, et al. Mapping of five new putative anion transporter genes 76. Safieddine S, El-Amraoui A, Petit C. The auditory hair cell
in human and characterization of SLC26A6, a candidate gene for ribbon synapse: From assembly to function. Ann Rev Neurosc.
pancreatic anion exchanger. Genomics. 2000. 2012;35:509-528.
58. Dallos P, Fakler B. Prestin, a new type of motor protein. Nat Rev Mol 77. Fettiplace R. Hair cell transduction, tuning, and synaptic
Cell Biol. 2002:104-111. transmission in the mammalian cochlea. Compr Physiol.
59. Matsunaga T, Morimoto N. The auditory phenotype of children 2017:1197-1227.
harboring mutations in the prestin gene. Acta Oto-Laryngol. 78. Nouvian R, et al. Structure and function of the hair cell ribbon
2016;136(4):397-401. synapse. J Memb Biol. 2006;209(2-3):153-165.
60. Liberman MC, Gao J, He DZ, et al. Prestin is required for electromotility 79. Fuchs PA, Lauer AM. Efferent inhibition of the cochlea.
of the outer hair cell and for the cochlear amplifie. Nature. Cold Spring Harbor Perspectives in Medicine. Cold Spring Harbor
2002;419(6904):300-304. Laboratory Press, 2019;9(5).
61. Mellado Lagarde MM, et al. Prestin’s Role in Cochlear Frequency 80. Roux I, et al. Otoferlin, Defective in a Human Deafness Form,
Tuning and Transmission of Mechanical Responses to Neural Is Essential for Exocytosis at the Auditory Ribbon Synapse. Cell.
Excitation. Curr Biol. 2008;18(3):200-202. 2006;127(2):277-289.
62. Dallos P, et al. Prestin-Based Outer Hair Cell Motility Is Necessary for 81. Yasunaga S, et al. A mutation in OTOF, encoding otoferlin, a FER-
Mammalian Cochlear Amplification. Neuron. 2008;58(3):333-339. 1-like protein, causes DFNB9, a nonsyndromic form of deafness. Nat
63. Martin P, Hudspeth AJ. Active hair-bundle movements can amplify Genet. 1999;21(4):363-369.
a hair cell’s response to oscillatory mechanical stimuli. Proceedings 82. Starr A, et al. Auditory neuropathy. Brain. 1996;119(3):741-753.
© 2021. Thieme. All rights reserved.
Você também pode gostar
- Seminario 28Documento14 páginasSeminario 28May MouraAinda não há avaliações
- Fisiologia Da AudiçãoDocumento5 páginasFisiologia Da Audiçãoursulagalvao100% (3)
- Anatomofisiologia Da AudiçãoDocumento4 páginasAnatomofisiologia Da AudiçãoIasminAinda não há avaliações
- ANAÌ TOMOFISIO AUDICÌ AÌ O e VESTIBULAR 1 ANO FONODocumento27 páginasANAÌ TOMOFISIO AUDICÌ AÌ O e VESTIBULAR 1 ANO FONOisabella persiraAinda não há avaliações
- Fisiologia Básica Da AudiçãoDocumento5 páginasFisiologia Básica Da AudiçãoCaio AmaralAinda não há avaliações
- Aula 4 - Biofísica Da AudiçãoDocumento15 páginasAula 4 - Biofísica Da AudiçãoGustavo Sergio SancinettiAinda não há avaliações
- Trabalho de Biofísica Da Audição.Documento11 páginasTrabalho de Biofísica Da Audição.Ricardo ValérioAinda não há avaliações
- O Ouvido HumanoDocumento19 páginasO Ouvido HumanoEdmilson ManganoteAinda não há avaliações
- Audição PDFDocumento8 páginasAudição PDFalinferAinda não há avaliações
- Aula 02 - Audicao 01 Correto 2020Documento56 páginasAula 02 - Audicao 01 Correto 2020Bio AndersonAinda não há avaliações
- Sistema Nervoso Sensorial Audição & EquilíbrioDocumento78 páginasSistema Nervoso Sensorial Audição & EquilíbrioRodrigo de Oliveira ReisAinda não há avaliações
- Biofisica Dos SistemasDocumento6 páginasBiofisica Dos SistemasCédrick CunhaAinda não há avaliações
- BIOFISICA Da AudiçãoDocumento13 páginasBIOFISICA Da AudiçãoandressafgaAinda não há avaliações
- Biofísica Da AudiçãoDocumento4 páginasBiofísica Da AudiçãoIvanPedroAinda não há avaliações
- Trabalho Fisiologia Luiza RevisadoDocumento7 páginasTrabalho Fisiologia Luiza RevisadoLuíza DidonéAinda não há avaliações
- Fechamento Problema 1Documento10 páginasFechamento Problema 1Wellington SouzaAinda não há avaliações
- Biofisica Da AudiçãoDocumento23 páginasBiofisica Da AudiçãoNicole CameloAinda não há avaliações
- Física Acústica para FonoaudiologiaDocumento8 páginasFísica Acústica para FonoaudiologiafilipecwaAinda não há avaliações
- Biofisica Da AudiçãoDocumento55 páginasBiofisica Da AudiçãoBio AndersonAinda não há avaliações
- Ruido1Documento41 páginasRuido1Tiago FerreiraAinda não há avaliações
- Resumo Fisiologia AudiçãoDocumento2 páginasResumo Fisiologia AudiçãovivianAinda não há avaliações
- Audicao 3Documento301 páginasAudicao 3Alice FerminoAinda não há avaliações
- Atributos Do Som e Sua Deteção Pelo Ser HumanoDocumento3 páginasAtributos Do Som e Sua Deteção Pelo Ser HumanoDaniel Neves DiasAinda não há avaliações
- Resumo - Bases Físicas Da Fonação, Audição e PsicoacústicaDocumento8 páginasResumo - Bases Físicas Da Fonação, Audição e PsicoacústicaLeonardo Brum100% (1)
- Anatomia Fisiológica Da AudiçãoDocumento3 páginasAnatomia Fisiológica Da AudiçãoGlória MamédioAinda não há avaliações
- A Física Na Audição PDFDocumento20 páginasA Física Na Audição PDFFernando José PeixotoAinda não há avaliações
- Aula 4 - AudiologiaDocumento25 páginasAula 4 - Audiologiamoovecase100% (1)
- Apresentação Biofísica OkDocumento21 páginasApresentação Biofísica OkTony NogueiraAinda não há avaliações
- Os Sons Do MundoDocumento24 páginasOs Sons Do MundoSantosAinda não há avaliações
- 1568920804-1552968663-Fisio Neuro Sistema Auditivo PDFDocumento9 páginas1568920804-1552968663-Fisio Neuro Sistema Auditivo PDFANA MARGARIDA MARGARIDAAinda não há avaliações
- Anatomia Da Orelha e OtitesDocumento20 páginasAnatomia Da Orelha e OtitesGuilhermeAinda não há avaliações
- Estudo Dirigido BIOFISICADocumento3 páginasEstudo Dirigido BIOFISICASaiaka IngridAinda não há avaliações
- Otorrinolaringologia - Resumo CompletoDocumento87 páginasOtorrinolaringologia - Resumo CompletoJulyana Lunardi Mousquer100% (4)
- Resumo - Som e AudiçãoDocumento20 páginasResumo - Som e AudiçãoFelipeAinda não há avaliações
- Aula 4 Audiologia PDFDocumento25 páginasAula 4 Audiologia PDFAndréa GarciaAinda não há avaliações
- AudiçãoDocumento3 páginasAudiçãoAlexAinda não há avaliações
- Apostila Sonoplastia PDFDocumento36 páginasApostila Sonoplastia PDFVladimir Borges100% (1)
- APRESENTAÇÃO BIOFÍSICA DA AUDIÇÃO 2 PeríodoDocumento13 páginasAPRESENTAÇÃO BIOFÍSICA DA AUDIÇÃO 2 PeríodoMarcos SilvaAinda não há avaliações
- Fisiologia - Roteiro 2 - Módulo 8Documento19 páginasFisiologia - Roteiro 2 - Módulo 8Ana Livia CamolesiAinda não há avaliações
- Spav Lista01Documento8 páginasSpav Lista01Natan Dos SantosAinda não há avaliações
- Aula Pratica - Sistema Auditivo e Vestibular - 2024Documento4 páginasAula Pratica - Sistema Auditivo e Vestibular - 2024vivisaggiomoAinda não há avaliações
- Resumo Sobre Anatomia Da OrelhaDocumento1 páginaResumo Sobre Anatomia Da OrelhaAna Karina MelendezAinda não há avaliações
- Acústica para Salas de Músicas PDFDocumento11 páginasAcústica para Salas de Músicas PDFEderAinda não há avaliações
- 7 - Sistemas VibrantesDocumento6 páginas7 - Sistemas VibrantesAzeitona Raphael GomesAinda não há avaliações
- Resumo P3Documento21 páginasResumo P3Sandro Natal DanielAinda não há avaliações
- Resumo PsicoacusticaDocumento8 páginasResumo PsicoacusticaIsabella Claudino FernandesAinda não há avaliações
- 2 - AudiçãoDocumento9 páginas2 - AudiçãobeatrizAinda não há avaliações
- Efeitos de Ruido e Vibrações No Homem PDFDocumento7 páginasEfeitos de Ruido e Vibrações No Homem PDFHugo Ferreira MooreAinda não há avaliações
- Fisiologia Da AudiçãoDocumento11 páginasFisiologia Da AudiçãoDione MendesAinda não há avaliações
- 1 - Anatomia e Fisiologia Do Órgao Da Audição e Do EquilíbrioDocumento14 páginas1 - Anatomia e Fisiologia Do Órgao Da Audição e Do EquilíbrioAnny AndradeAinda não há avaliações
- Aula Anatomia e Fisiologia Da AudiçãoDocumento36 páginasAula Anatomia e Fisiologia Da AudiçãoLiliane Pan100% (1)
- Apo - HeltonDocumento9 páginasApo - HeltonMaria LauraAinda não há avaliações
- 77988d90-de39-11ea-b706-0fc8bf1e2202Documento6 páginas77988d90-de39-11ea-b706-0fc8bf1e2202eng civil Paulo Vitor MedeirosAinda não há avaliações
- Aula 4Documento7 páginasAula 4ets2girlasmrAinda não há avaliações
- Bases Da AudiçãoDocumento10 páginasBases Da AudiçãoGraziela AzevedoAinda não há avaliações
- IMITANCIOMETRIA - ResumidaDocumento31 páginasIMITANCIOMETRIA - ResumidaGabrielle LimaAinda não há avaliações
- Biofisica Da AudicaoDocumento29 páginasBiofisica Da AudicaoAna Katarine Pordeus LisboaAinda não há avaliações
- A linguagem das crianças surdas: Guia para familiares e educadoresNo EverandA linguagem das crianças surdas: Guia para familiares e educadoresAinda não há avaliações
- PORTARIA #140, DE 21 DE MARÇO DE 2022 - PORTARIA #140, DE 21 DE MARÇO DE 2022 - DOU - Imprensa NacionalDocumento94 páginasPORTARIA #140, DE 21 DE MARÇO DE 2022 - PORTARIA #140, DE 21 DE MARÇO DE 2022 - DOU - Imprensa NacionalJoão VieiraAinda não há avaliações
- Silo - Tips Sistema de Osmose Reversa BaumerDocumento12 páginasSilo - Tips Sistema de Osmose Reversa BaumerFrancisco MartinezAinda não há avaliações
- MANUAL DE INSTRUCAO CTSI Caminhao Transbordo Santa IzabelDocumento16 páginasMANUAL DE INSTRUCAO CTSI Caminhao Transbordo Santa IzabelZeka Borgato0% (1)
- Tabela de Componentes Elétricos CompressoresDocumento12 páginasTabela de Componentes Elétricos CompressoresSergio PinheiroAinda não há avaliações
- Apostila Eletrotécnica. e Acionamentos - ResistoresDocumento17 páginasApostila Eletrotécnica. e Acionamentos - Resistoresvictor hugoAinda não há avaliações
- Circulo de Mohr Ua IDocumento18 páginasCirculo de Mohr Ua IEder RibeiroAinda não há avaliações
- Causas Da Flutuação de TensãoDocumento57 páginasCausas Da Flutuação de TensãoJorge AzevedoAinda não há avaliações
- Ondas Sismicas DescontinuidadesDocumento20 páginasOndas Sismicas DescontinuidadesBery UrrutiaAinda não há avaliações
- MONOGRAFIA EstudoImplantaçãoSistemaDocumento65 páginasMONOGRAFIA EstudoImplantaçãoSistemaCarlos ChavesAinda não há avaliações
- Experimento 5Documento3 páginasExperimento 5João FelipeAinda não há avaliações
- EJ - RESUMO NAV PP PlanejamentoDocumento5 páginasEJ - RESUMO NAV PP PlanejamentoFabio Sant Ana de OliveiraAinda não há avaliações
- 12ano F 1 1 5 Movimentos de Corpos Sujeitos A LigacoesDocumento6 páginas12ano F 1 1 5 Movimentos de Corpos Sujeitos A Ligacoesmafaldatavares31Ainda não há avaliações
- Roteiro 10 - Sintese - Cu - Glicinato - Espectroscopia UV-Vis - Versaonova - 2023 - QuiÌ Mica e Engenharia - 02032023-2Documento13 páginasRoteiro 10 - Sintese - Cu - Glicinato - Espectroscopia UV-Vis - Versaonova - 2023 - QuiÌ Mica e Engenharia - 02032023-2Victor RodriguesAinda não há avaliações
- Avaliação de Matemática - 3 Unidade - Expressões AlgébricasDocumento3 páginasAvaliação de Matemática - 3 Unidade - Expressões AlgébricasJuliana AraújoAinda não há avaliações
- Bantam Serralheiro 110220 VDocumento16 páginasBantam Serralheiro 110220 VDiegoAinda não há avaliações
- Descrição de Itens Vig Ambiental CorrigidaDocumento6 páginasDescrição de Itens Vig Ambiental CorrigidaAlbert HenriqueAinda não há avaliações
- Fa1 ProfDocumento6 páginasFa1 ProfcaiogabrielAinda não há avaliações
- Ceperj 2012 Cedae RJ Engenheiro Eletricista ProvaDocumento8 páginasCeperj 2012 Cedae RJ Engenheiro Eletricista Provafelipealves13Ainda não há avaliações
- Relatório Completo 2Documento12 páginasRelatório Completo 2Camila Sciena100% (1)
- Resumo - Concretos EspeciaisDocumento7 páginasResumo - Concretos EspeciaisKaue Kawashita100% (1)
- 2013 ESCAVADEIRA HIDRAULICA JCB JS130LC 1108-FE015 Folheto Tecnico JS130LC - Janeiro - 2013Documento8 páginas2013 ESCAVADEIRA HIDRAULICA JCB JS130LC 1108-FE015 Folheto Tecnico JS130LC - Janeiro - 2013Cassius BenitesAinda não há avaliações
- Exerc Oscilacoes OndasDocumento9 páginasExerc Oscilacoes OndasGislayne de Lima SantosAinda não há avaliações
- NBR 6489 Solo - Prova de Carga Estática em Fundação DiretaDocumento15 páginasNBR 6489 Solo - Prova de Carga Estática em Fundação DiretaTássia NetoAinda não há avaliações
- Ficha de PreparacaoDocumento5 páginasFicha de PreparacaoNelson0% (1)
- NBR 15575 - Original PDFDocumento388 páginasNBR 15575 - Original PDFDouglasAinda não há avaliações
- Aterramento em Canteiro de Obras - Instruções Básicas - Vinicius AyrãoDocumento15 páginasAterramento em Canteiro de Obras - Instruções Básicas - Vinicius AyrãoMauricio Baeta Neves FilhoAinda não há avaliações
- EX FQA715 F1 2022 V1 - NetDocumento16 páginasEX FQA715 F1 2022 V1 - Netdrecas12842Ainda não há avaliações
- PGF rp10213 2002-06Documento26 páginasPGF rp10213 2002-06Emerson André Bueno VolpeAinda não há avaliações
- RS8020Documento3 páginasRS8020Alan MarcioAinda não há avaliações
- NBR5356 1Documento103 páginasNBR5356 1Bia AlvesAinda não há avaliações