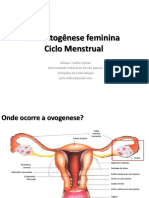
Você também pode gostar
- Biologi 3Documento32 páginasBiologi 3Jamiles Andrade100% (1)
- Gametogênese HumanaDocumento20 páginasGametogênese Humanajoaopedroolk2129Ainda não há avaliações
- GametogeneseDocumento29 páginasGametogeneseFabio Rodrigues de MeloAinda não há avaliações
- Aula 1 - 2 - Gametogenese Feminina e MasculinaDocumento62 páginasAula 1 - 2 - Gametogenese Feminina e MasculinaBruno Machado BertassoliAinda não há avaliações
- Formação de gametas e meiose reduz cromossomosDocumento15 páginasFormação de gametas e meiose reduz cromossomosludmillarangelAinda não há avaliações
- Bio 12Documento10 páginasBio 12ana carolinaAinda não há avaliações
- GametogeneseDocumento26 páginasGametogeneseHonda Civic 2006 seminovoAinda não há avaliações
- EMBRIOLOGIADocumento6 páginasEMBRIOLOGIAMaria Clara MardeganAinda não há avaliações
- Gametogênese: formação dos gametas masculinos e femininosDocumento14 páginasGametogênese: formação dos gametas masculinos e femininosMylena karolyne Souza silvaAinda não há avaliações
- Gametogênese e Ciclo MenstrualDocumento30 páginasGametogênese e Ciclo MenstrualYam VictorAinda não há avaliações
- Aula 1 Módulo C - Fecundação e 1 Semana Do DesenvolvimentoDocumento39 páginasAula 1 Módulo C - Fecundação e 1 Semana Do DesenvolvimentoLarissa FardinAinda não há avaliações
- Espermatogênese e oogêneseDocumento2 páginasEspermatogênese e oogêneseAldesio Francisco da Costa FilhoAinda não há avaliações
- Relatório 3 BDDocumento4 páginasRelatório 3 BDSavio SitoeAinda não há avaliações
- Embrio P1Documento49 páginasEmbrio P1issehclineuzieg09Ainda não há avaliações
- EMBRIOLOGIADocumento22 páginasEMBRIOLOGIAAna LídiaAinda não há avaliações
- OvogeneseDocumento39 páginasOvogenesemedicina7x1Ainda não há avaliações
- 3 - Gametogênese e Fertilização HumanaDocumento37 páginas3 - Gametogênese e Fertilização HumanamaribserraAinda não há avaliações
- e7378f44-3ae2-4c48-a848-b80adf530dc9Documento21 páginase7378f44-3ae2-4c48-a848-b80adf530dc9Larissa MoraisAinda não há avaliações
- Gametogénese e Regulação HormonalDocumento53 páginasGametogénese e Regulação HormonalMaria SantosAinda não há avaliações
- A descoberta da célula e os principais componentesDocumento26 páginasA descoberta da célula e os principais componentesMARIA CLEMILDA SILVAAinda não há avaliações
- Null 2Documento12 páginasNull 2adaoAinda não há avaliações
- Apuntes de EmbriologíaDocumento6 páginasApuntes de Embriologíacarlos castroAinda não há avaliações
- Sistema Rep. Humano e Gamet. - GametogêneseDocumento21 páginasSistema Rep. Humano e Gamet. - GametogênesePaulo HenriqueAinda não há avaliações
- Sistema ReprodutorDocumento37 páginasSistema ReprodutorSoares MaunatoAinda não há avaliações
- Embriologia IDocumento55 páginasEmbriologia IJuliaAinda não há avaliações
- GametogeneseDocumento22 páginasGametogeneselucas carvalhoAinda não há avaliações
- Resumo Modular 1Documento7 páginasResumo Modular 1Bruno Souza FonsecaAinda não há avaliações
- Revisão EmbriologiaDocumento6 páginasRevisão EmbriologiaGiovanna AlbuquerqueAinda não há avaliações
- Embriologia Humana - Gametogenese - Aula 1Documento88 páginasEmbriologia Humana - Gametogenese - Aula 1Jéssica AlmeidaAinda não há avaliações
- Ovogênese e formação de folículosDocumento28 páginasOvogênese e formação de folículosFelipe RomãoAinda não há avaliações
- Biologia 1 - GametogêneseDocumento11 páginasBiologia 1 - GametogêneseVIVI LOUSANOAinda não há avaliações
- Apostila - EmbriologiaDocumento96 páginasApostila - EmbriologiaGuilhermeSantosAinda não há avaliações
- Introdução À Genética Capítulo 1Documento28 páginasIntrodução À Genética Capítulo 1kelstonmilianiAinda não há avaliações
- 2 Gametogênese FemininaDocumento11 páginas2 Gametogênese FemininaIhury JhonsonAinda não há avaliações
- Oogenese Artigo CienciaelementarDocumento2 páginasOogenese Artigo Cienciaelementardiana BaptistaAinda não há avaliações
- Ovogenese e EspermatogeneseDocumento73 páginasOvogenese e EspermatogeneseDjavan0% (1)
- Gamotenese - BiologiaDocumento2 páginasGamotenese - Biologiaalexsandra santosAinda não há avaliações
- Questionario Av2 2021Documento5 páginasQuestionario Av2 2021Letycia Fernandes100% (1)
- OvogeneseDocumento28 páginasOvogeneseNulciene Freitas100% (1)
- Ciclo Menstrual (Atualizado)Documento20 páginasCiclo Menstrual (Atualizado)Mariane Patricio CostaAinda não há avaliações
- O ciclo celularDocumento31 páginasO ciclo celularsilrogerioAinda não há avaliações
- Resumo Da Aula Pratica 3Documento6 páginasResumo Da Aula Pratica 3Naira ClementinaAinda não há avaliações
- Embriologia MédicaDocumento68 páginasEmbriologia Médicaanarodriguesmed28Ainda não há avaliações
- Espermatogenese e Oogenese IdoxDocumento15 páginasEspermatogenese e Oogenese IdoxIdolêncio NhangaveAinda não há avaliações
- EmbriologiaDocumento8 páginasEmbriologiaThiago Soldatelli BritoAinda não há avaliações
- Ovogénese em Ascaris: As fases da maturaçãoDocumento6 páginasOvogénese em Ascaris: As fases da maturaçãoNaira ClementinaAinda não há avaliações
- 1 Aula Prof: MirianDocumento80 páginas1 Aula Prof: MirianNerfanda MaiaAinda não há avaliações
- Introdução à Genética: Herança Biológica e Transmissão de CaracterísticasDocumento28 páginasIntrodução à Genética: Herança Biológica e Transmissão de CaracterísticasLudimila Oliveira100% (1)
- Estudo Dirigido Embrio IDocumento4 páginasEstudo Dirigido Embrio IdaienescamposAinda não há avaliações
- Aula 1 - Embrio - 2019Documento56 páginasAula 1 - Embrio - 2019Helena FláviaAinda não há avaliações
- Curso 161075 Aula 03 Grifado 0482Documento46 páginasCurso 161075 Aula 03 Grifado 0482DergreneAinda não há avaliações
- Desenvolvimento EmbrionárioDocumento101 páginasDesenvolvimento Embrionárioallan felipeAinda não há avaliações
- Anatomia Do Sistema Reprodutor GametogéneseDocumento52 páginasAnatomia Do Sistema Reprodutor GametogéneseBruna CostaAinda não há avaliações
- Oogénese e estrutura dos ováriosDocumento18 páginasOogénese e estrutura dos ováriostatianaAinda não há avaliações
- Gametogênese Humana, HistologiaDocumento22 páginasGametogênese Humana, HistologiaDavi PeronicaAinda não há avaliações
- Fisiologia Da Reprodução HumanaDocumento23 páginasFisiologia Da Reprodução HumanaPaulaAinda não há avaliações
- EMBRIOLOGIA Primeira Semana - Curso Med 2023/1 NicDocumento6 páginasEMBRIOLOGIA Primeira Semana - Curso Med 2023/1 NicNicolas Iulius Lussari BrisonAinda não há avaliações
- Desenvolvimento embrionário e anexosDocumento29 páginasDesenvolvimento embrionário e anexosRita RodriguesAinda não há avaliações
- Aula 1 - Embriologia - Gametogênese e FecundaçãoDocumento38 páginasAula 1 - Embriologia - Gametogênese e FecundaçãoJulia BragaAinda não há avaliações
- Prova 2 Citologia Áleff ADocumento5 páginasProva 2 Citologia Áleff AÁleff Allan Oliveira BorgesAinda não há avaliações
- Xaden Pov 16Documento8 páginasXaden Pov 16Thais BertoldiAinda não há avaliações
- Histologia do tecido nervoso e estruturas cerebraisDocumento8 páginasHistologia do tecido nervoso e estruturas cerebraisAdrielleAinda não há avaliações
- Walter Moreira Maia Júnior - Tese PPGRN 2003.Documento268 páginasWalter Moreira Maia Júnior - Tese PPGRN 2003.Jonatas PauloAinda não há avaliações
- Edicao32 - Lorena TiemyDocumento8 páginasEdicao32 - Lorena TiemyKelly Neves BooneAinda não há avaliações
- Material de Apoio - Cinética de Crescimento CelularDocumento42 páginasMaterial de Apoio - Cinética de Crescimento CelularJameson ToledoAinda não há avaliações
- Mod 3 P07Documento15 páginasMod 3 P07Rayane rodrigues dos santosAinda não há avaliações
- 5 Sociobiologia ControversiaDocumento5 páginas5 Sociobiologia ControversiaEuro VillaAinda não há avaliações
- E Book Disciplina 13 Atuacao em Equipe Multiprofissional e Intersetorialidade PPTX 2 1669301584Documento46 páginasE Book Disciplina 13 Atuacao em Equipe Multiprofissional e Intersetorialidade PPTX 2 1669301584Jandiara MagalhãesAinda não há avaliações
- Aula3 - Introdução À Sistemática Vegetal 2Documento34 páginasAula3 - Introdução À Sistemática Vegetal 2Matheus Augsuto VeigaAinda não há avaliações
- Alem Do Azul - Jussara LealDocumento618 páginasAlem Do Azul - Jussara LealEdilane GonçalvesAinda não há avaliações
- Transformação Do Músculo em Carne - Rigor MortisDocumento48 páginasTransformação Do Músculo em Carne - Rigor MortisDaniela Maria100% (1)
- DNA na prática: extraindo e explorando o DNADocumento6 páginasDNA na prática: extraindo e explorando o DNARAISSAAinda não há avaliações
- 2° Prova de FisiologiaDocumento5 páginas2° Prova de FisiologiaPaulla Nathalia Oliveira SantanaAinda não há avaliações
- Operações de Multiplicação e Divisão Significados: NúmerosDocumento19 páginasOperações de Multiplicação e Divisão Significados: NúmerosLucieleDeHolandaAinda não há avaliações
- Neuro CiênciaDocumento6 páginasNeuro CiênciafcoAinda não há avaliações
- Roteiro Da Prova de Bases Prática IDocumento5 páginasRoteiro Da Prova de Bases Prática ISugar MelonAinda não há avaliações
- Divisão Celular e CrescimentoDocumento40 páginasDivisão Celular e CrescimentotrilibiteAinda não há avaliações
- Fisiologia Humana: Líquidos CorporaisDocumento2 páginasFisiologia Humana: Líquidos CorporaisWendell BuenoAinda não há avaliações
- Relatorio Aula Pratica Analises Clinicas Reticulocitos - EricaDocumento6 páginasRelatorio Aula Pratica Analises Clinicas Reticulocitos - Ericadado.sp9Ainda não há avaliações
- Relatorio 5 .......................Documento2 páginasRelatorio 5 .......................rodrigoerodrigues1971Ainda não há avaliações
- Bioética e desenvolvimento sustentávelDocumento14 páginasBioética e desenvolvimento sustentávelBruna OlíviaAinda não há avaliações
- Relatorio Biologia MolecularDocumento11 páginasRelatorio Biologia Moleculardelricianee clementina da silvaAinda não há avaliações
- Gene Interaction Heritability in PeppersDocumento6 páginasGene Interaction Heritability in PeppersAline MissioAinda não há avaliações
- ES foi o Estado que mais investiu em 2021Documento135 páginasES foi o Estado que mais investiu em 2021Carlos Magno Novaes AzevedoAinda não há avaliações
- Epitélios, Conjuntivo e Adiposo: Características e FunçõesDocumento51 páginasEpitélios, Conjuntivo e Adiposo: Características e FunçõesMarcelle Gonçalves de MirandaAinda não há avaliações
- Caça Palavras Sobre EcologiaDocumento1 páginaCaça Palavras Sobre EcologiaJanayna GentilinAinda não há avaliações
- Sapiens 2024Documento22 páginasSapiens 2024ThaywanderleyAinda não há avaliações
- Higiene ocupacional em atividades industriaisDocumento268 páginasHigiene ocupacional em atividades industriaisRonaldo RodrgiesAinda não há avaliações
- 1000 Most Common Portuguese WordsDocumento22 páginas1000 Most Common Portuguese WordsAlexandru TacheAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Serviços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesNo EverandServiços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesAinda não há avaliações
- Parasitologia 1: Helmintos de Interesse MédicoNo EverandParasitologia 1: Helmintos de Interesse MédicoAinda não há avaliações
- Educação ambiental: A formação do sujeito ecológicoNo EverandEducação ambiental: A formação do sujeito ecológicoAinda não há avaliações
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Fundamentos Em Cirurgia OralNo EverandFundamentos Em Cirurgia OralNota: 5 de 5 estrelas5/5 (1)
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- A clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNo EverandA clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNota: 3.5 de 5 estrelas3.5/5 (3)
- Fisiologia Cardiorrespiratória Ensino SuperiorNo EverandFisiologia Cardiorrespiratória Ensino SuperiorAinda não há avaliações
- Topônimos no Sul da Bahia: nominações dos municípios originados da capitania hereditária de São Jorge dos Ilheus (1950 a 1960)No EverandTopônimos no Sul da Bahia: nominações dos municípios originados da capitania hereditária de São Jorge dos Ilheus (1950 a 1960)Nota: 5 de 5 estrelas5/5 (1)
- Concentração: Torne-se mais consciente e economize tempoNo EverandConcentração: Torne-se mais consciente e economize tempoNota: 4.5 de 5 estrelas4.5/5 (3)