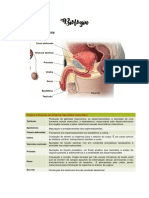
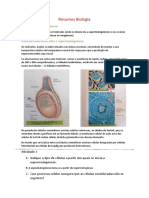
Você também pode gostar
- Bio 220 4Documento6 páginasBio 220 4Gabrielly PaivaAinda não há avaliações
- Estudo Dirigido Embrio IDocumento4 páginasEstudo Dirigido Embrio IdaienescamposAinda não há avaliações
- Espermatogênese:: Espermatócitos PrimáriosDocumento10 páginasEspermatogênese:: Espermatócitos PrimáriosLuciene FriasAinda não há avaliações
- Embrio P1Documento49 páginasEmbrio P1issehclineuzieg09Ainda não há avaliações
- Da Fecundação Ao NascimentoDocumento9 páginasDa Fecundação Ao NascimentoFaBeAnAinda não há avaliações
- GametogeneseDocumento6 páginasGametogeneseLetícia BoniniAinda não há avaliações
- Primeira Semana - Pré ClivagemDocumento3 páginasPrimeira Semana - Pré ClivagemHellen KathleenAinda não há avaliações
- Resumos Fecundação - Processo Notocordal PDFDocumento11 páginasResumos Fecundação - Processo Notocordal PDFmarambaiaditoAinda não há avaliações
- EmbriologiaDocumento6 páginasEmbriologiaLetícia BoniniAinda não há avaliações
- GametogeneseDocumento14 páginasGametogeneseBelo LopezAinda não há avaliações
- 1 Semana Do Desenvolvimento EmbrionárioDocumento16 páginas1 Semana Do Desenvolvimento EmbrionárioGláucia LaísAinda não há avaliações
- Revisão EmbriologiaDocumento21 páginasRevisão EmbriologialailaribeirocostaAinda não há avaliações
- Regulação Hormonal Da OogéneseDocumento4 páginasRegulação Hormonal Da OogéneseMatilde EstevesAinda não há avaliações
- Embriologia - Primeira SemanaDocumento7 páginasEmbriologia - Primeira SemanaMaria Clara TabosaAinda não há avaliações
- Resumo Da Primeira Semana Do Desenvolvimento WordDocumento8 páginasResumo Da Primeira Semana Do Desenvolvimento WordMaria Esthela Bull LooseAinda não há avaliações
- Primeira Semana Do DesenvolvimentoDocumento11 páginasPrimeira Semana Do DesenvolvimentoLone GhostAinda não há avaliações
- Módulo de Concepção-Tutoria 3-Fertilização e BlastocistoDocumento15 páginasMódulo de Concepção-Tutoria 3-Fertilização e BlastocistoMatheus UrsiAinda não há avaliações
- BiologiaDocumento16 páginasBiologiaJoel NogueiraAinda não há avaliações
- Embriologia Humana - Parte 1Documento30 páginasEmbriologia Humana - Parte 1Yam VictorAinda não há avaliações
- ReproduçãoDocumento4 páginasReproduçãoMiguelAinda não há avaliações
- EMBRIOLOGIADocumento6 páginasEMBRIOLOGIAMaria Clara MardeganAinda não há avaliações
- Questionario Av2 2021Documento5 páginasQuestionario Av2 2021Letycia Fernandes100% (1)
- Resumo de EmbriologiaDocumento12 páginasResumo de EmbriologiaDaniela silvaAinda não há avaliações
- Matéria de Embriologia Comparada - Biologia Até NEURULAÇÃODocumento34 páginasMatéria de Embriologia Comparada - Biologia Até NEURULAÇÃODérik FilipeAinda não há avaliações
- Biologia 12º Ano - Resumo - Reprodução HumanaDocumento24 páginasBiologia 12º Ano - Resumo - Reprodução HumanaStephane Azevedo100% (21)
- Reprodução e Manipulação Da FertilidadeDocumento10 páginasReprodução e Manipulação Da FertilidadebeatrizromaoAinda não há avaliações
- Gametogénese e Regulação HormonalDocumento53 páginasGametogénese e Regulação HormonalMaria SantosAinda não há avaliações
- Resumos Biologia ClassromDocumento6 páginasResumos Biologia ClassromLil FerreiraAinda não há avaliações
- Relatório Da Aula Experimental. Estrutura Das Gónadas e Gâmetas Humanos I Sérgio Tomás Valentim (Final) PDFDocumento14 páginasRelatório Da Aula Experimental. Estrutura Das Gónadas e Gâmetas Humanos I Sérgio Tomás Valentim (Final) PDFJoao MAchado100% (1)
- Preparando Gametas e Iniciando Os Eventos EmbrionáriosDocumento15 páginasPreparando Gametas e Iniciando Os Eventos EmbrionáriosalcantarabrennaAinda não há avaliações
- Resumo de Embriologia PDFDocumento17 páginasResumo de Embriologia PDFDanielaAlvesAinda não há avaliações
- Gametogênese Humana, HistologiaDocumento22 páginasGametogênese Humana, HistologiaDavi PeronicaAinda não há avaliações
- Historiologia e EmbriologiaDocumento2 páginasHistoriologia e EmbriologiaRayne CamposAinda não há avaliações
- 2 - Segunda Semana Do Desenvolvimento HumanoDocumento5 páginas2 - Segunda Semana Do Desenvolvimento HumanoKarlla RaphaellaAinda não há avaliações
- Relatorio Biologia 2022 FINALLDocumento17 páginasRelatorio Biologia 2022 FINALLMariana Simoes100% (1)
- Primeiras Semanas EMBRIODocumento3 páginasPrimeiras Semanas EMBRIOColégio FochAinda não há avaliações
- Resumo EmbrioDocumento21 páginasResumo EmbriochiclebluejmAinda não há avaliações
- Resumo ''Gametogenese Humana'' FPMDocumento7 páginasResumo ''Gametogenese Humana'' FPMJoanaAinda não há avaliações
- S1 - AULA 1 & 2 - Gametogênese, Fecun., Implan., 1, 2 e 3ra SemanaDocumento12 páginasS1 - AULA 1 & 2 - Gametogênese, Fecun., Implan., 1, 2 e 3ra SemanaCayê AlvesAinda não há avaliações
- EMBRIOLOGIADocumento16 páginasEMBRIOLOGIAFernanda Rocha DinizAinda não há avaliações
- Resumo - Reprodução Humana e Manioulação Da FertilidadeDocumento23 páginasResumo - Reprodução Humana e Manioulação Da FertilidadeMartaRuasAinda não há avaliações
- Escola Iqra - Nazia Trabalho de Biologia (Guardado Automaticamente)Documento12 páginasEscola Iqra - Nazia Trabalho de Biologia (Guardado Automaticamente)Cássimo LucianoAinda não há avaliações
- Gametogênese HumanaDocumento20 páginasGametogênese Humanajoaopedroolk2129Ainda não há avaliações
- SP3 - Onde Os Fracos Não Tem VezDocumento8 páginasSP3 - Onde Os Fracos Não Tem VezRenan VillacaAinda não há avaliações
- Embriologia ResumoDocumento45 páginasEmbriologia ResumoJuliana SilvaAinda não há avaliações
- Rel Bio 12Documento6 páginasRel Bio 12Luísa Braz TeixeiraAinda não há avaliações
- Biologia 12º Ano ReproduçãoDocumento10 páginasBiologia 12º Ano ReproduçãoRaquelAinda não há avaliações
- GametogeneseDocumento22 páginasGametogeneselucas carvalhoAinda não há avaliações
- 1° e 2° Semana Do Desenvolvimento EmbrionárioDocumento9 páginas1° e 2° Semana Do Desenvolvimento EmbrionárioAmandaAinda não há avaliações
- Relatório 1Documento14 páginasRelatório 1Alexandra BalçaAinda não há avaliações
- Resumos Unidade 1Documento9 páginasResumos Unidade 1Mariana SofiaAinda não há avaliações
- FecundaçãoDocumento3 páginasFecundaçãoRoberth Alexandro Soares de OliveiraAinda não há avaliações
- Resumo AV1 e AV2 - Embriologia e HistologiaDocumento11 páginasResumo AV1 e AV2 - Embriologia e HistologiaLuciene BarretoAinda não há avaliações
- EMBRIOLOGIADocumento22 páginasEMBRIOLOGIAAna LídiaAinda não há avaliações
- PPT3 - Fecundação, Desenvolvimento Embrionário e AnexosDocumento29 páginasPPT3 - Fecundação, Desenvolvimento Embrionário e AnexosRita RodriguesAinda não há avaliações
- Apg Desenvolvimento Embrionario e FetalDocumento12 páginasApg Desenvolvimento Embrionario e FetalGabriela RochaAinda não há avaliações
- P1M1 - AmenorreiaDocumento19 páginasP1M1 - AmenorreiaHemily RodriguesAinda não há avaliações
- Gametogênese e Desenvolvimento EmbrionárioDocumento19 páginasGametogênese e Desenvolvimento EmbrionárioAna Paula Do NascimentoAinda não há avaliações
- UntitledDocumento25 páginasUntitledBruno Souza FonsecaAinda não há avaliações
- Atividade IntegradoraDocumento2 páginasAtividade IntegradoraBruno Souza FonsecaAinda não há avaliações
- Educação Popular em Saúde IDocumento2 páginasEducação Popular em Saúde IBruno Souza FonsecaAinda não há avaliações
- Piesc IDocumento1 páginaPiesc IBruno Souza FonsecaAinda não há avaliações
- Portfolio-Hospital-Brasileiro-unopar-COPIA-sqhbrk 2Documento13 páginasPortfolio-Hospital-Brasileiro-unopar-COPIA-sqhbrk 2Bruno Souza FonsecaAinda não há avaliações
- ELETROSTATICADocumento37 páginasELETROSTATICAGilmar FerrettiAinda não há avaliações
- Catalogo Materiais Macea PDFDocumento3 páginasCatalogo Materiais Macea PDFTruno LaenderAinda não há avaliações
- SimuladoDocumento125 páginasSimuladosantos kAinda não há avaliações
- Prova Ensino T Cnico Concomitante Ou Subsequente e T Cnico Integrado Ao Ensino M DioDocumento12 páginasProva Ensino T Cnico Concomitante Ou Subsequente e T Cnico Integrado Ao Ensino M Dio46f97zw8cnAinda não há avaliações
- Roteiros de Estudo ItaimeDocumento3 páginasRoteiros de Estudo ItaimeMARCOS GABRIEL CATARINO FERREIRAAinda não há avaliações
- Indaclean Sabonete Liquido Perolado Erva Doce 0119 FispqDocumento4 páginasIndaclean Sabonete Liquido Perolado Erva Doce 0119 FispqAgnaldoAinda não há avaliações
- Catálogo GasoterapiaDocumento12 páginasCatálogo Gasoterapiaronaldobandeira0% (1)
- NomeclaturaDocumento10 páginasNomeclaturaIlla OliveiraAinda não há avaliações
- Tratamento Do Efluente Madeireiro Por Processo Foto-Fenton: Treatment of Timber Effluent by Photo-Fenton ProcessDocumento9 páginasTratamento Do Efluente Madeireiro Por Processo Foto-Fenton: Treatment of Timber Effluent by Photo-Fenton ProcessrocahAinda não há avaliações
- Furação Brocas - DormerDocumento12 páginasFuração Brocas - DormerDalton MarinoAinda não há avaliações
- Relatorio Derteminação Da Capacidade Térmica Do CalorímetroDocumento9 páginasRelatorio Derteminação Da Capacidade Térmica Do Calorímetromekino_silva100% (2)
- Relações de Maxwell - ResumoDocumento10 páginasRelações de Maxwell - ResumoJanaina LeitinhoAinda não há avaliações
- Questoes Quimica 9anoDocumento3 páginasQuestoes Quimica 9anopedroaugusto1313Ainda não há avaliações
- Apostila e Matéria PrimaDocumento17 páginasApostila e Matéria PrimasebiancoAinda não há avaliações
- 53 Amida 90 - Ficha TecnicaDocumento2 páginas53 Amida 90 - Ficha TecnicaCarolina TarifAinda não há avaliações
- REM Termografia e UltravioletaDocumento44 páginasREM Termografia e UltravioletaGustavo FernandesAinda não há avaliações
- C3 CursoE Biologia B 20aulasDocumento12 páginasC3 CursoE Biologia B 20aulasHancock86Ainda não há avaliações
- Vinificacoes EspeciaisDocumento41 páginasVinificacoes EspeciaisSilvino SilAinda não há avaliações
- Aula 7 - Cinetica Enzimatica PDFDocumento31 páginasAula 7 - Cinetica Enzimatica PDFViviane BorelaAinda não há avaliações
- Document - Onl - Formulas de Grafiatotexturatintasselador e Massa CorridaDocumento48 páginasDocument - Onl - Formulas de Grafiatotexturatintasselador e Massa CorridaAlison Andrade100% (1)
- Relatório 3 - Determinação de ProteínasDocumento3 páginasRelatório 3 - Determinação de ProteínasLiana FlorAinda não há avaliações
- Trabalho de MatecoDocumento2 páginasTrabalho de MatecoMiguel OliveiraAinda não há avaliações
- Ficha Tecnica Penecrete MortarDocumento4 páginasFicha Tecnica Penecrete MortarJoão DiasAinda não há avaliações
- Relatório Cloro AtivoDocumento16 páginasRelatório Cloro AtivoGAndrade TanyaAinda não há avaliações
- Relatório Invertase - Cinética e InibiçãoDocumento16 páginasRelatório Invertase - Cinética e InibiçãoLetícia M. Sgobi100% (1)
- Manual de Estabilidade PH de Ativos 2 - Ed PDFDocumento235 páginasManual de Estabilidade PH de Ativos 2 - Ed PDFSandroluciozontaAinda não há avaliações
- Maquinas TermicasDocumento74 páginasMaquinas Termicasjbastos12049079Ainda não há avaliações
- Aula 5 - Transferência de CalorDocumento45 páginasAula 5 - Transferência de CalorEvaldo MarquesAinda não há avaliações
- FISPQ - Cimento Portland NASSAU-1Documento2 páginasFISPQ - Cimento Portland NASSAU-1bruna brunaAinda não há avaliações