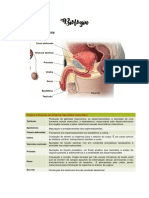
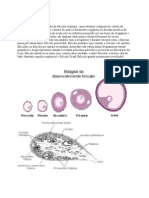
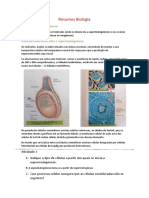
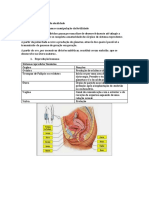
Você também pode gostar
- Desenvolvimento embrionário na 1a semana: fecundação, clivagem e implantaçãoDocumento16 páginasDesenvolvimento embrionário na 1a semana: fecundação, clivagem e implantaçãoGláucia LaísAinda não há avaliações
- Espermatogênese:: Espermatócitos PrimáriosDocumento10 páginasEspermatogênese:: Espermatócitos PrimáriosLuciene FriasAinda não há avaliações
- Gametogênese, Fecundação e As 3 Primeiras Semanas Do Desenvolvimento Embrionário PDFDocumento13 páginasGametogênese, Fecundação e As 3 Primeiras Semanas Do Desenvolvimento Embrionário PDFAbielAinda não há avaliações
- Estudo Dirigido Embrio IDocumento4 páginasEstudo Dirigido Embrio IdaienescamposAinda não há avaliações
- GametogeneseDocumento6 páginasGametogeneseLetícia BoniniAinda não há avaliações
- BiologiaDocumento16 páginasBiologiaJoel NogueiraAinda não há avaliações
- EmbriologiaDocumento6 páginasEmbriologiaLetícia BoniniAinda não há avaliações
- Primeira Semana - Pré ClivagemDocumento3 páginasPrimeira Semana - Pré ClivagemHellen KathleenAinda não há avaliações
- Resumo de Embriologia PDFDocumento17 páginasResumo de Embriologia PDFDanielaAlvesAinda não há avaliações
- Resumo Modular 1Documento7 páginasResumo Modular 1Bruno Souza FonsecaAinda não há avaliações
- Embriologia - Primeira SemanaDocumento7 páginasEmbriologia - Primeira SemanaMaria Clara TabosaAinda não há avaliações
- GametogeneseDocumento14 páginasGametogeneseBelo LopezAinda não há avaliações
- Processo de Formação dos GametasDocumento45 páginasProcesso de Formação dos GametasJuliana SilvaAinda não há avaliações
- Primeira Semana Desenvolvimento EmbrionárioDocumento11 páginasPrimeira Semana Desenvolvimento EmbrionárioLone GhostAinda não há avaliações
- Matéria de Embriologia Comparada - Biologia Até NEURULAÇÃODocumento34 páginasMatéria de Embriologia Comparada - Biologia Até NEURULAÇÃODérik FilipeAinda não há avaliações
- Resumo de EmbriologiaDocumento15 páginasResumo de Embriologiaapi-3700467100% (6)
- Embriologia 1-3 Semana ResumoDocumento13 páginasEmbriologia 1-3 Semana ResumoRachel MonroeAinda não há avaliações
- Formação de gametas femininos e masculinosDocumento18 páginasFormação de gametas femininos e masculinoswelkleyAinda não há avaliações
- Processos de Gametogênese e FecundaçãoDocumento12 páginasProcessos de Gametogênese e FecundaçãoCayê AlvesAinda não há avaliações
- Da Fecundação Ao NascimentoDocumento9 páginasDa Fecundação Ao NascimentoFaBeAnAinda não há avaliações
- Resumo de EmbriologiaDocumento12 páginasResumo de EmbriologiaDaniela silvaAinda não há avaliações
- Gametogénese e Regulação HormonalDocumento53 páginasGametogénese e Regulação HormonalMaria SantosAinda não há avaliações
- EMBRIOLOGIADocumento6 páginasEMBRIOLOGIAMaria Clara MardeganAinda não há avaliações
- Tutoria Problema I - Fechamento Ii 1. Descrever As Fases Da FecundaçãoDocumento12 páginasTutoria Problema I - Fechamento Ii 1. Descrever As Fases Da FecundaçãoBárbara Cristal Oliveira AlmeidaAinda não há avaliações
- Reprodução e manipulação da fertilidadeDocumento10 páginasReprodução e manipulação da fertilidadebeatrizromaoAinda não há avaliações
- Gametogênese HumanaDocumento20 páginasGametogênese Humanajoaopedroolk2129Ainda não há avaliações
- Resumos Fecundação - Processo Notocordal PDFDocumento11 páginasResumos Fecundação - Processo Notocordal PDFmarambaiaditoAinda não há avaliações
- Biologia 1 - GametogêneseDocumento11 páginasBiologia 1 - GametogêneseVIVI LOUSANOAinda não há avaliações
- Embrio P1Documento49 páginasEmbrio P1issehclineuzieg09Ainda não há avaliações
- Fertilização, clivagem e implantação inicialDocumento8 páginasFertilização, clivagem e implantação inicialMaria Esthela Bull LooseAinda não há avaliações
- Biologia - 12ºanoDocumento7 páginasBiologia - 12ºanopedro luísAinda não há avaliações
- Gametogênese, Fecundação e NidaçãoDocumento30 páginasGametogênese, Fecundação e NidaçãoHelena Flávia0% (1)
- Fecundação: início do desenvolvimento humanoDocumento3 páginasFecundação: início do desenvolvimento humanoRoberth Alexandro Soares de OliveiraAinda não há avaliações
- Gametogênese e FecundaçãoDocumento19 páginasGametogênese e Fecundaçãoalline alvesAinda não há avaliações
- Desenvolvimento Humano nas Primeiras Duas SemanasDocumento9 páginasDesenvolvimento Humano nas Primeiras Duas SemanasAmandaAinda não há avaliações
- FOLICULOGÊNESE Ntit (Recovered)Documento8 páginasFOLICULOGÊNESE Ntit (Recovered)Simone Salvador BoeneAinda não há avaliações
- Resumo ''Gametogenese Humana'' FPMDocumento7 páginasResumo ''Gametogenese Humana'' FPMJoanaAinda não há avaliações
- Módulo de Concepção-Tutoria 3-Fertilização e BlastocistoDocumento15 páginasMódulo de Concepção-Tutoria 3-Fertilização e BlastocistoMatheus UrsiAinda não há avaliações
- Resumo - Reprodução Humana e Manioulação Da FertilidadeDocumento23 páginasResumo - Reprodução Humana e Manioulação Da FertilidadeMartaRuasAinda não há avaliações
- Historiologia e EmbriologiaDocumento2 páginasHistoriologia e EmbriologiaRayne CamposAinda não há avaliações
- Processo de fecundação e desenvolvimento embrionárioDocumento12 páginasProcesso de fecundação e desenvolvimento embrionárioGabriela RochaAinda não há avaliações
- Embriologia - TranscriçãoDocumento21 páginasEmbriologia - TranscriçãoHygor OliveiraAinda não há avaliações
- Formação dos Gametas e Concepção HumanaDocumento43 páginasFormação dos Gametas e Concepção Humanathiago melo dos anjosAinda não há avaliações
- Biologia 12.º AnoDocumento65 páginasBiologia 12.º AnoPaula Amado100% (1)
- Gametogênese e Ciclo MenstrualDocumento30 páginasGametogênese e Ciclo MenstrualYam VictorAinda não há avaliações
- Gametogênese e Desenvolvimento EmbrionárioDocumento19 páginasGametogênese e Desenvolvimento EmbrionárioAna Paula Do NascimentoAinda não há avaliações
- GametogeneseDocumento22 páginasGametogeneselucas carvalhoAinda não há avaliações
- Resumo Biologia Gónadas e gametogéneseDocumento6 páginasResumo Biologia Gónadas e gametogéneseLil FerreiraAinda não há avaliações
- 12º BiologiaDocumento12 páginas12º BiologiaMiriam CruzAinda não há avaliações
- ReproduçãoDocumento4 páginasReproduçãoMiguelAinda não há avaliações
- Resumo de Embriologia - Prova 1Documento14 páginasResumo de Embriologia - Prova 1Maria Clara DolceAinda não há avaliações
- Gametogéneses e Ovogénese Trabalho BiologiaDocumento12 páginasGametogéneses e Ovogénese Trabalho BiologiaCássimo LucianoAinda não há avaliações
- Biologia 12ºDocumento71 páginasBiologia 12ºKatia RodriguesAinda não há avaliações
- Biologia 12º Ano ReproduçãoDocumento10 páginasBiologia 12º Ano ReproduçãoRaquelAinda não há avaliações
- Resumo Embriologia MooreDocumento45 páginasResumo Embriologia MooreRaquel Ferreira100% (2)
- Relatório Aula Prática MeioseDocumento9 páginasRelatório Aula Prática MeioseRenas Pereira75% (4)
- Embriologia Ensino Superior Resumo PDFDocumento41 páginasEmbriologia Ensino Superior Resumo PDFAry Junior100% (1)
- Regulação Hormonal Da OogéneseDocumento4 páginasRegulação Hormonal Da OogéneseMatilde EstevesAinda não há avaliações