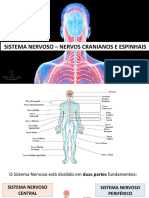
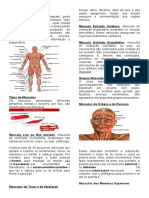
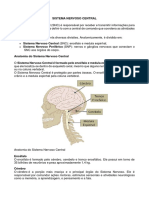
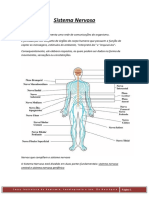
Você também pode gostar
- Anatomia e Fisiologia Humana 2Documento30 páginasAnatomia e Fisiologia Humana 2Daniele LealAinda não há avaliações
- Origem Das CidadesDocumento23 páginasOrigem Das Cidadesthiago aquinoAinda não há avaliações
- Trabalho para Gracieta-1Documento5 páginasTrabalho para Gracieta-1Dominic GittinAinda não há avaliações
- 5-Reação de Polimerização em CadeiaDocumento4 páginas5-Reação de Polimerização em CadeiaLaiana TrindadeAinda não há avaliações
- Fisiologia Do Sistema Nervoso Somático e Sistema Nervoso VisceralDocumento53 páginasFisiologia Do Sistema Nervoso Somático e Sistema Nervoso VisceralRafael BitencourtAinda não há avaliações
- Resumo Neuro N3Documento26 páginasResumo Neuro N3Paulo BarrosAinda não há avaliações
- Aula 06 - Contração Muscular - Corpo HumanoDocumento38 páginasAula 06 - Contração Muscular - Corpo HumanoLiliane MeloAinda não há avaliações
- Aula de Neurofisiologia Aspectos Basicos Dos MovimentosDocumento32 páginasAula de Neurofisiologia Aspectos Basicos Dos MovimentosMichelle Monaliza100% (1)
- Sistema Nervoso SomáticoDocumento11 páginasSistema Nervoso SomáticoorinelAinda não há avaliações
- Fisiologia para Fisioterapia - Aula 5Documento23 páginasFisiologia para Fisioterapia - Aula 5julliagabriela2009Ainda não há avaliações
- Fisiologiadamotricidade 110118100510 Phpapp01 PDFDocumento5 páginasFisiologiadamotricidade 110118100510 Phpapp01 PDFGabriela TeixeiraAinda não há avaliações
- CAPÍTULO 11.fisiologiaDocumento5 páginasCAPÍTULO 11.fisiologiaNatalia CanabravaAinda não há avaliações
- HistologiaDocumento15 páginasHistologiaGeorgia TortelliAinda não há avaliações
- Funções Motoras Do Sistema Nervoso CentralDocumento16 páginasFunções Motoras Do Sistema Nervoso CentralMonitoriadefisio100% (3)
- Slide 7 e 8Documento66 páginasSlide 7 e 8sidioushenriAinda não há avaliações
- Sistemas Nervoso e Musculoesquelético - Reação Do Corpo em Relação Aos Aspectos ErgonômicoDocumento42 páginasSistemas Nervoso e Musculoesquelético - Reação Do Corpo em Relação Aos Aspectos ErgonômicoMichel RobertoAinda não há avaliações
- Aula 4 - Sistema Nervoso CentralDocumento29 páginasAula 4 - Sistema Nervoso CentralKeile Anne Silva100% (1)
- Sistema Nervoso - Beatriz CordeiroDocumento4 páginasSistema Nervoso - Beatriz CordeiroBeatriz de Oliveira CordeiroAinda não há avaliações
- 2 - Comando Nervoso e MovimentosDocumento38 páginas2 - Comando Nervoso e MovimentosPaloma DrummondAinda não há avaliações
- Introdução Ao Sistema NervosoDocumento4 páginasIntrodução Ao Sistema Nervososaidefelix mariaAinda não há avaliações
- Sistema NervosoDocumento16 páginasSistema NervosoFrancielle SampaioAinda não há avaliações
- SNA Tortora InformaçõesDocumento14 páginasSNA Tortora InformaçõesAna lauraAinda não há avaliações
- Complexo Escolar Privado 18 de Novembro KimanguitaDocumento11 páginasComplexo Escolar Privado 18 de Novembro Kimanguitajosuemikael11Ainda não há avaliações
- 12 Aula Sistema NervosoDocumento25 páginas12 Aula Sistema NervosoShopee suporteAinda não há avaliações
- M4P4Documento7 páginasM4P4Hemily RodriguesAinda não há avaliações
- Apostila - Sistema Nervoso PDFDocumento13 páginasApostila - Sistema Nervoso PDF.Ainda não há avaliações
- Sistem Nervoso 4. ExercíciosDocumento4 páginasSistem Nervoso 4. ExercíciosAna AntiqueiraAinda não há avaliações
- Sistema NervosoDocumento7 páginasSistema Nervososandra ribeiroAinda não há avaliações
- Neurofisiologia - Prova 3Documento25 páginasNeurofisiologia - Prova 3fernanda.alves18Ainda não há avaliações
- Estudo Dirigido NeuroanatomiaDocumento8 páginasEstudo Dirigido NeuroanatomiaAriel Gomes100% (1)
- Controle - Motricidade or Nishida UnespDocumento27 páginasControle - Motricidade or Nishida UnespRegis AssisAinda não há avaliações
- Resumo - Sistema NervosoDocumento8 páginasResumo - Sistema NervosoThayane MoreiraAinda não há avaliações
- Sistema Nervoso Autônomo (SNA)Documento11 páginasSistema Nervoso Autônomo (SNA)EDEN GUADALUPE SIQUEIRAAinda não há avaliações
- Sistema NervosoDocumento41 páginasSistema NervosoJoão LucasAinda não há avaliações
- Sistema NeuroHormonalDocumento31 páginasSistema NeuroHormonalmargarida-pimentel-3439Ainda não há avaliações
- Aula Aminoácidos AtualDocumento9 páginasAula Aminoácidos AtualGabryel LeiteAinda não há avaliações
- Sistema NervosoDocumento3 páginasSistema NervosohombedjoAinda não há avaliações
- Alunos Com Deficiência Física NeuromotoraDocumento9 páginasAlunos Com Deficiência Física NeuromotoraEllen Aparecida MarangoniAinda não há avaliações
- Fisiologia para Fisioterapia - Aula 6Documento19 páginasFisiologia para Fisioterapia - Aula 6julliagabriela2009Ainda não há avaliações
- Sistema NervosoDocumento14 páginasSistema NervosoEricka RodriguesAinda não há avaliações
- Sistemas para ImprimirDocumento10 páginasSistemas para ImprimirGLEICILENE SILVAAinda não há avaliações
- Sistema NervosoDocumento37 páginasSistema NervosoLara StefaneAinda não há avaliações
- Sistema Nervoso CentralDocumento3 páginasSistema Nervoso CentralAna Paula Oliveira0% (1)
- O Sistema NervosoDocumento3 páginasO Sistema NervosoGéssica GutierresAinda não há avaliações
- O Sistema Nervoso PeriféricoDocumento4 páginasO Sistema Nervoso Periféricoyohan GomesAinda não há avaliações
- Aula 2 - NeurofisiologiaDocumento38 páginasAula 2 - NeurofisiologiaLeandro LimaAinda não há avaliações
- Aula 6 m02 Sistema NervosoDocumento7 páginasAula 6 m02 Sistema NervosolarissaaugustobioAinda não há avaliações
- Capítulo 54 - Funções Motoras Da Medula Espinhal - Os Reflexos Espinhais - 4 PáginasDocumento4 páginasCapítulo 54 - Funções Motoras Da Medula Espinhal - Os Reflexos Espinhais - 4 PáginasandreamokadaAinda não há avaliações
- Resumo Sistema NervosoDocumento3 páginasResumo Sistema NervosoguilhermeAinda não há avaliações
- Fisiologia Humana SNC SNP ImpressãoDocumento48 páginasFisiologia Humana SNC SNP ImpressãoLuciano FreitasAinda não há avaliações
- O Sistema Nervoso PeriféricoDocumento6 páginasO Sistema Nervoso PeriféricoKélita SilvaAinda não há avaliações
- Material de Apoio - Sistema NervosoDocumento4 páginasMaterial de Apoio - Sistema NervosoJuliana PereiraAinda não há avaliações
- Sistema NervosoDocumento11 páginasSistema NervosoAlcides EmidioAinda não há avaliações
- Sistema NervosoDocumento3 páginasSistema Nervosotailson santos da silvaAinda não há avaliações
- Sistema NervosoDocumento59 páginasSistema NervosoMaiara De Paula Santana FerreiraAinda não há avaliações
- Relatório Do Sistema NervosoDocumento12 páginasRelatório Do Sistema Nervosomirandaninteres100% (1)
- 6 Ano Caderno4 Semana 4 o Sistema Nervoso 1 PDFDocumento7 páginas6 Ano Caderno4 Semana 4 o Sistema Nervoso 1 PDFWill SAinda não há avaliações
- Sistema Nervoso Parte 2Documento37 páginasSistema Nervoso Parte 2DionatanAinda não há avaliações
- Sistema NervosoDocumento5 páginasSistema NervosoCassy Mandri InstrutoraAinda não há avaliações
- Aparelho Visual + Auditivo + CoordenaçãoDocumento5 páginasAparelho Visual + Auditivo + CoordenaçãoInês FariaAinda não há avaliações
- Sistema NervosoDocumento3 páginasSistema NervosoLili Ribeiro100% (1)
- Temática - Ciências da Vida - Relação e CoordenaçãoNo EverandTemática - Ciências da Vida - Relação e CoordenaçãoAinda não há avaliações
- Turma - 2021 - Plano de Pormenor de Nassapo-3Documento27 páginasTurma - 2021 - Plano de Pormenor de Nassapo-3AmaroAinda não há avaliações
- Projeto 3 Idade - Qualidade de VidaDocumento12 páginasProjeto 3 Idade - Qualidade de VidaClauDinho Moraes0% (1)
- ERP Apostila SGI PDFDocumento56 páginasERP Apostila SGI PDFFilipe PereiraAinda não há avaliações
- André Malina e Ângela Azevedo - O Esporte Pode Tudo - o Livro Síntese Do Intelectual Vitor Marinho de OliveiraDocumento5 páginasAndré Malina e Ângela Azevedo - O Esporte Pode Tudo - o Livro Síntese Do Intelectual Vitor Marinho de OliveiraOdraudeIttereipAinda não há avaliações
- Livro - Material de Produção IndustrialDocumento298 páginasLivro - Material de Produção IndustrialSilvandiraRodrigues0% (1)
- Atividade - MauraDocumento4 páginasAtividade - MauraDonilio Vinicius LisboaAinda não há avaliações
- A Revolucao Nacional de 1926Documento27 páginasA Revolucao Nacional de 1926Sónia FernandesAinda não há avaliações
- A Inclusão Escolar de Uma CriançaDocumento13 páginasA Inclusão Escolar de Uma CriançaConrado PlothowAinda não há avaliações
- Hot Oil SprayDocumento3 páginasHot Oil SprayMárcio Vieira SilvaAinda não há avaliações
- Revolução IndustrialDocumento13 páginasRevolução IndustrialMarcondes100% (1)
- Trabalho II Simposio Internacional Bibliotecas DigitaisDocumento50 páginasTrabalho II Simposio Internacional Bibliotecas Digitaismari_daviAinda não há avaliações
- Manual de Instrucoes de Instalacao e Montagem Valvula de Retencao Atuada r1 9785fceb25Documento7 páginasManual de Instrucoes de Instalacao e Montagem Valvula de Retencao Atuada r1 9785fceb25André Gasparino SchmidtAinda não há avaliações
- OProgramadeeducaopr Escolardeacordocomaconcepopedaggicadobem AventuradoEdmundoBojanowski Rozklad 2Documento66 páginasOProgramadeeducaopr Escolardeacordocomaconcepopedaggicadobem AventuradoEdmundoBojanowski Rozklad 2Abel Kambinda JambaAinda não há avaliações
- 14 Projeto Fluxo de CaixaDocumento13 páginas14 Projeto Fluxo de CaixaAmanda Dreger PeresAinda não há avaliações
- CaligramasDocumento13 páginasCaligramasEmmanuelAinda não há avaliações
- Air Bike Semana 1Documento7 páginasAir Bike Semana 1Victor Carvalho NogueiraAinda não há avaliações
- O Povo Tuxá e o Dzubukuá. Ricardo Menezes BarbosaDocumento2 páginasO Povo Tuxá e o Dzubukuá. Ricardo Menezes BarbosaRicardo MenezesAinda não há avaliações
- Payment InstructionsDocumento2 páginasPayment InstructionsThiago Souza da SilvaAinda não há avaliações
- Atividade Diagnóstica 02 - EmpreendedorismoDocumento2 páginasAtividade Diagnóstica 02 - EmpreendedorismoEdjailson OliveiraAinda não há avaliações
- Familiarização Do TBMDocumento33 páginasFamiliarização Do TBMMarcos MartinsAinda não há avaliações
- Orientacoes PET EFTI Volume IV 2020Documento50 páginasOrientacoes PET EFTI Volume IV 2020Cleo SousaAinda não há avaliações
- Manual de Reiki Essencial IDocumento14 páginasManual de Reiki Essencial IO Sítio de Neptuno100% (1)
- Material de ConsultaDocumento7 páginasMaterial de ConsultaÉlida AlvesAinda não há avaliações
- Relatório Torre de - 1Documento5 páginasRelatório Torre de - 1Ricardo NunesAinda não há avaliações
- UFCD ReflexãoDocumento2 páginasUFCD ReflexãokimmikAinda não há avaliações
- Atividades de FisicaDocumento4 páginasAtividades de Fisicaleda_professoraAinda não há avaliações
- Módulo 4 Do Curso de Limpeza Protecção e Magia AngelicalDocumento19 páginasMódulo 4 Do Curso de Limpeza Protecção e Magia AngelicalPaulo SantosAinda não há avaliações