Você também pode gostar
- Modelo de Relatorio de NecropsiaDocumento2 páginasModelo de Relatorio de NecropsiaVet_arquivos79% (14)
- 8-Exame Andrológico Do TouroDocumento52 páginas8-Exame Andrológico Do Tourolucaskkk123Ainda não há avaliações
- Exame Ginecológico em Pequeno AnimaisDocumento7 páginasExame Ginecológico em Pequeno AnimaisGrazi HonorioAinda não há avaliações
- 4-MANEJO SANITÁRIO VACAS LEITEIRAS DefDocumento12 páginas4-MANEJO SANITÁRIO VACAS LEITEIRAS DefMarciliana Davantel100% (1)
- Palestra Parto e Distocias em Cães - Quando IntervirDocumento28 páginasPalestra Parto e Distocias em Cães - Quando IntervirFernanda Trigo de AzevedoAinda não há avaliações
- Gestão em Pequenas Propriedades LeiteirasDocumento14 páginasGestão em Pequenas Propriedades LeiteirasPaulo GodoiAinda não há avaliações
- Alguns Pontos Importantes em Exames Clínicos de AnimaisDocumento4 páginasAlguns Pontos Importantes em Exames Clínicos de AnimaisVet_arquivosAinda não há avaliações
- A Importância Do Médico Veterinário Na Saúde PúblicaDocumento47 páginasA Importância Do Médico Veterinário Na Saúde PúblicaVet_arquivos100% (3)
- SuinoculturaDocumento27 páginasSuinoculturaVet_arquivos100% (3)
- Farmacologia - Cos de Uso Frequente Na Cli Nica VeterinariaDocumento24 páginasFarmacologia - Cos de Uso Frequente Na Cli Nica Veterinariaapi-3814824100% (10)
- Relatório - Aula Prática de CápsulasDocumento10 páginasRelatório - Aula Prática de CápsulasnunosajAinda não há avaliações
- PSICOLOGIA APLICADA À IMPLANTODONTIA E ProteseDocumento40 páginasPSICOLOGIA APLICADA À IMPLANTODONTIA E ProteseJuliana Esteche67% (3)
- Femur Joelho PernaDocumento57 páginasFemur Joelho PernaBira Vianna100% (1)
- Transtorno de EliminaçãoDocumento41 páginasTranstorno de EliminaçãoAlladin&Eventos AO33% (3)
- Retenção Placentária Na ÉguaDocumento79 páginasRetenção Placentária Na ÉguaPRISCILLA DAYANNE CHAGAS SOUZAAinda não há avaliações
- Fisiovet - FemeaDocumento15 páginasFisiovet - FemeaWerner AlbuquerqueAinda não há avaliações
- Transferencia de EmbrioesDocumento43 páginasTransferencia de Embrioesfanni_lalaAinda não há avaliações
- Afecções Do Sistema Locomotor de BovinosDocumento26 páginasAfecções Do Sistema Locomotor de Bovinosrodrigoferrazza1662Ainda não há avaliações
- Genoma BovinoDocumento18 páginasGenoma BovinoFlávio LimaAinda não há avaliações
- Anatomia Do Aparelho Reprodutor Do MachoDocumento26 páginasAnatomia Do Aparelho Reprodutor Do MachoAlexandre Silva de AlmeidaAinda não há avaliações
- Aspectos Da Reprodução Equina: Inseminação Artificial E Tranferência de Embrião: Revisão de LiteraturaDocumento23 páginasAspectos Da Reprodução Equina: Inseminação Artificial E Tranferência de Embrião: Revisão de LiteraturaAdryano CarvalhoAinda não há avaliações
- 1583408338GUIA TCNICO - Passos para o Sucesso Na Inseminao Artificial PDFDocumento46 páginas1583408338GUIA TCNICO - Passos para o Sucesso Na Inseminao Artificial PDFRubens BorgesAinda não há avaliações
- Abdome Agudo Equino Controle Da DorDocumento9 páginasAbdome Agudo Equino Controle Da DorNim WildAinda não há avaliações
- Gomes Et Al., 2015. Exame Andrologico em BovinosDocumento6 páginasGomes Et Al., 2015. Exame Andrologico em Bovinosrodrigoferrazza1662Ainda não há avaliações
- Manual de Exame Ginecológico Da VacaDocumento5 páginasManual de Exame Ginecológico Da VacamarciogomesgvAinda não há avaliações
- Iatf, Te e Fiv em Bufalos PDFDocumento14 páginasIatf, Te e Fiv em Bufalos PDFdanimedeiirosAinda não há avaliações
- Fisiologia Da Reprodução em Gatas 2011Documento71 páginasFisiologia Da Reprodução em Gatas 2011silviacrusco100% (1)
- Manejo Reprodutivo em Bovinos de CorteDocumento57 páginasManejo Reprodutivo em Bovinos de CorteMatheus GurgelAinda não há avaliações
- Degeneração TesticularDocumento38 páginasDegeneração TesticularCarolina Berkman100% (4)
- Aula8 Desenho de PrimersDocumento59 páginasAula8 Desenho de PrimersCarlianeAinda não há avaliações
- Peritonite em EquinosDocumento16 páginasPeritonite em EquinosDaizy QueirozAinda não há avaliações
- Periodo Seco Vacas LeiteirasDocumento4 páginasPeriodo Seco Vacas LeiteirasCybervetAinda não há avaliações
- Controle Zootecnico em Bovinos de CorteDocumento3 páginasControle Zootecnico em Bovinos de CorteGelson LodiAinda não há avaliações
- Atendimento Emergencial de FraturaDocumento10 páginasAtendimento Emergencial de FraturaAlana SoaresAinda não há avaliações
- Ciclo Estral BovinosDocumento33 páginasCiclo Estral BovinosswaggerleeAinda não há avaliações
- Sistema Reprodutor Dos EquinosDocumento33 páginasSistema Reprodutor Dos EquinosNeide100% (3)
- 2 Fertilidade e Obstetricia em Bovinos - NoakesDocumento146 páginas2 Fertilidade e Obstetricia em Bovinos - NoakesSílvia CarvalhoAinda não há avaliações
- Clinica de GrandesDocumento10 páginasClinica de GrandesEsaias SilvaAinda não há avaliações
- Bovinos - Morfofisiologia e Microbiologia Do Rúmen PDFDocumento29 páginasBovinos - Morfofisiologia e Microbiologia Do Rúmen PDFJunior OliveiraAinda não há avaliações
- Inseminação Artificial em SuinosDocumento90 páginasInseminação Artificial em SuinosJúlio Cesar PereiraAinda não há avaliações
- Tabela Exigencias Alimentos BovinosDocumento24 páginasTabela Exigencias Alimentos Bovinosmara coutoAinda não há avaliações
- Retenção de Placenta em VacasDocumento7 páginasRetenção de Placenta em VacasVet_arquivos100% (1)
- As Prostaglandinas Na ReproduçãoDocumento26 páginasAs Prostaglandinas Na ReproduçãoVet_arquivos100% (2)
- Conceitos Basicos de Nutricao de Ruminantes - Fabio PrudencioDocumento101 páginasConceitos Basicos de Nutricao de Ruminantes - Fabio PrudencioGiovani PastreAinda não há avaliações
- Digestão e Absorção de Produtos 2020.2Documento61 páginasDigestão e Absorção de Produtos 2020.2Tata PetroffAinda não há avaliações
- Slides - PartoDocumento34 páginasSlides - PartoLaura SchulzAinda não há avaliações
- Heterose e HeterozigoseDocumento25 páginasHeterose e Heterozigoselucildalima100% (1)
- Fisiologia Veterinária II - Ciclos Reprodutivos Bovinos e OvinosDocumento25 páginasFisiologia Veterinária II - Ciclos Reprodutivos Bovinos e OvinosAlbertoGonçalvesEvangelistaAinda não há avaliações
- Cuidado Neonato EquinoDocumento3 páginasCuidado Neonato Equinosemiologia tardeAinda não há avaliações
- Helmintos de Ruminantes e EquinosDocumento52 páginasHelmintos de Ruminantes e EquinosJonas Machado CunicoAinda não há avaliações
- Afecções Da Glândula Mamária Dos Ruminantes IDocumento27 páginasAfecções Da Glândula Mamária Dos Ruminantes IAmanda AliceAinda não há avaliações
- Aula SarnasDocumento59 páginasAula SarnasGedeao Candido CamisaAinda não há avaliações
- Manual de Rotulagem de POA CIPOA SISP Jan 2019Documento43 páginasManual de Rotulagem de POA CIPOA SISP Jan 2019Siluana Benvindo Ferreira100% (1)
- Aspectos Do Ciclo Estral Da GataDocumento41 páginasAspectos Do Ciclo Estral Da GataJoao Paulo TeixeiraAinda não há avaliações
- Afecções Do Sistema Genitourinário em RuminantesDocumento83 páginasAfecções Do Sistema Genitourinário em RuminantesAmanda AliceAinda não há avaliações
- O Manejo Sanitário e Sua Importância No Novo Contexto Do Agronegócio Da Produção de Pecuária de CorteDocumento5 páginasO Manejo Sanitário e Sua Importância No Novo Contexto Do Agronegócio Da Produção de Pecuária de CorteAugusta RomanielloAinda não há avaliações
- Métodos Semiológicos de Exame Físico em RuminantesDocumento5 páginasMétodos Semiológicos de Exame Físico em RuminantesVet_arquivos100% (3)
- Doce de LeiteDocumento3 páginasDoce de LeiteÉrika PatriciaAinda não há avaliações
- Equívocos Na Cólica EquinaDocumento7 páginasEquívocos Na Cólica EquinaRaíssa GalegoAinda não há avaliações
- Manual Bea (Dgav)Documento112 páginasManual Bea (Dgav)Lara TeixeiraAinda não há avaliações
- Tã Cnica Cirã Rgica Celiotomia e Digestã Rio de Grandes AnimaisDocumento91 páginasTã Cnica Cirã Rgica Celiotomia e Digestã Rio de Grandes AnimaisTata PetroffAinda não há avaliações
- Brucelose e Tuberculose Bovina 2 PDFDocumento98 páginasBrucelose e Tuberculose Bovina 2 PDFSimone BalianAinda não há avaliações
- 2 - Fisiologia Do Ciclo Estral Nas Especies Domesticas GabrielDocumento57 páginas2 - Fisiologia Do Ciclo Estral Nas Especies Domesticas GabrielHériton SilvaAinda não há avaliações
- Reproducao AvesDocumento11 páginasReproducao AvesbiancadrumondAinda não há avaliações
- Aula 04 - Fisiologia e Endocrinologia Da Reprodução Da FêmeaDocumento74 páginasAula 04 - Fisiologia e Endocrinologia Da Reprodução Da FêmeaHELIDA FERNANDES LEAOAinda não há avaliações
- Aula 9 Lactação Fisio II 2020Documento54 páginasAula 9 Lactação Fisio II 2020Betânia Glória CamposAinda não há avaliações
- Avaliação Ginecológica e Utilização Racional de Protocolos ReprodutivosDocumento26 páginasAvaliação Ginecológica e Utilização Racional de Protocolos ReprodutivosMarcus Vinicius Vieira GomesAinda não há avaliações
- Sistema Urinario 2012Documento72 páginasSistema Urinario 2012José GonçalvesAinda não há avaliações
- Controle Endócrino Do Equilíbrio Hidro-EletrolíticoDocumento18 páginasControle Endócrino Do Equilíbrio Hidro-EletrolíticoVet_arquivosAinda não há avaliações
- As Prostaglandinas Na ReproduçãoDocumento26 páginasAs Prostaglandinas Na ReproduçãoVet_arquivos100% (2)
- Sarna Sarcoptica em Caes PDFDocumento5 páginasSarna Sarcoptica em Caes PDFMJoanaDuarteAinda não há avaliações
- Choque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaDocumento22 páginasChoque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaVet_arquivosAinda não há avaliações
- Curso VendedorDocumento138 páginasCurso VendedorVet_arquivosAinda não há avaliações
- Timpanismo em RuminAntesDocumento6 páginasTimpanismo em RuminAntesVet_arquivos100% (1)
- Causa e Tratamento Do Garrotilho em EquinosDocumento5 páginasCausa e Tratamento Do Garrotilho em EquinosVet_arquivos100% (1)
- Noções Básicas de FarmacologiaDocumento16 páginasNoções Básicas de FarmacologiaVet_arquivos100% (5)
- Doenças Respiratórias Dos BovinosDocumento6 páginasDoenças Respiratórias Dos BovinosVet_arquivosAinda não há avaliações
- Carbúnculo HemáticoDocumento5 páginasCarbúnculo HemáticoVet_arquivos100% (1)
- Atrofia e NecroseDocumento7 páginasAtrofia e NecroseVet_arquivosAinda não há avaliações
- Retenção de Placenta em VacasDocumento7 páginasRetenção de Placenta em VacasVet_arquivos100% (1)
- Carbúnculo Sintomático em BovinoDocumento5 páginasCarbúnculo Sintomático em BovinoVet_arquivosAinda não há avaliações
- Digestão de Celulose Nos VertebradosDocumento2 páginasDigestão de Celulose Nos VertebradosVet_arquivosAinda não há avaliações
- Terapêutica de Aves SilvestresDocumento6 páginasTerapêutica de Aves SilvestresVet_arquivosAinda não há avaliações
- Inovações Técnológicas Na BioclimatologiaDocumento24 páginasInovações Técnológicas Na BioclimatologiaVet_arquivos100% (1)
- Ciclo Estral Nos AnimaisDocumento15 páginasCiclo Estral Nos AnimaisVet_arquivos57% (7)
- Métodos Semiológicos de Exame Físico em RuminantesDocumento5 páginasMétodos Semiológicos de Exame Físico em RuminantesVet_arquivos100% (3)
- Raças de CãesDocumento98 páginasRaças de CãesVet_arquivos100% (4)
- Nutrição de Animais HospitalizadosDocumento14 páginasNutrição de Animais HospitalizadosVet_arquivos100% (1)
- Prescrição Racional em Medicina VeterináriaDocumento10 páginasPrescrição Racional em Medicina VeterináriaVet_arquivosAinda não há avaliações
- Gestação e Parto em Cadelas Fisiologia, Diagnóstico de Gestação e Tratamento de DistociasDocumento9 páginasGestação e Parto em Cadelas Fisiologia, Diagnóstico de Gestação e Tratamento de DistociasVet_arquivosAinda não há avaliações
- Fatores Que Influenciam MastiteDocumento15 páginasFatores Que Influenciam MastiteVet_arquivosAinda não há avaliações
- Zoonoses e Doenças Emergentes Transmitidas Por Animais SilvestresDocumento7 páginasZoonoses e Doenças Emergentes Transmitidas Por Animais SilvestresVet_arquivos100% (1)
- SISTEMA CARDIOVASCULAR Novo-1Documento18 páginasSISTEMA CARDIOVASCULAR Novo-1EdsonAinda não há avaliações
- Prova Biologia (2018)Documento19 páginasProva Biologia (2018)carla soares100% (1)
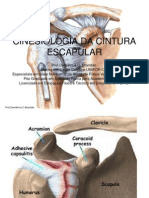
- 06 - Cinesiologia Da Cintura EscapularDocumento34 páginas06 - Cinesiologia Da Cintura EscapularJoão Paulo Ramos OliveiraAinda não há avaliações
- Fitoterapia Aplicada A EsteticaDocumento133 páginasFitoterapia Aplicada A EsteticaHans WandebergAinda não há avaliações
- AnatomoFisiologia Do Sistema DigestórioDocumento54 páginasAnatomoFisiologia Do Sistema DigestórioBreno de Souza MotaAinda não há avaliações
- Calcificação e Pigmentos - PatologiaDocumento21 páginasCalcificação e Pigmentos - PatologiaBianca BiancaAinda não há avaliações
- Resumo - Vascularização Cabeça e Pescoço - Documentos GoogleDocumento20 páginasResumo - Vascularização Cabeça e Pescoço - Documentos GoogleEstudos VitóriaAinda não há avaliações
- Apresentação 1 Introdução Aos Fungos ATUALIZADODocumento28 páginasApresentação 1 Introdução Aos Fungos ATUALIZADOJamille BritoAinda não há avaliações
- Apostila de Fisiologia - Maria EduardaDocumento64 páginasApostila de Fisiologia - Maria EduardaMIRIA PORTO LIMA100% (2)
- SENSAÇÃO, PERCEPÇÃO E CONSCIÊNCIA - MindMeister Mapa MentalDocumento1 páginaSENSAÇÃO, PERCEPÇÃO E CONSCIÊNCIA - MindMeister Mapa MentalLaura FernandaAinda não há avaliações
- Aula - 5Documento43 páginasAula - 5texugolokuAinda não há avaliações
- Apresentação de MetodologiaDocumento19 páginasApresentação de MetodologiaGuilherme Vaz MarquesAinda não há avaliações
- Ovos de Yoni - OvoterapiaDocumento6 páginasOvos de Yoni - Ovoterapiapriiskaf100% (2)
- RespiratórioDocumento34 páginasRespiratórioPlínio RodriguesAinda não há avaliações
- A Importância Dos AlongamentosDocumento15 páginasA Importância Dos AlongamentosPhilipe De Paiva SouzaAinda não há avaliações
- Lista de Exercicio de BioquimicaDocumento4 páginasLista de Exercicio de BioquimicaJenifer Sofia de SouzaAinda não há avaliações
- Teste Gosto-Intensidade Análise SensorialDocumento10 páginasTeste Gosto-Intensidade Análise SensorialErica Carvalho100% (1)
- OSSOSDocumento3 páginasOSSOSAnonymous wTC8bDaLAinda não há avaliações
- Medicina AlternativaDocumento4 páginasMedicina AlternativaELazaroAinda não há avaliações
- Mapa Mental 2Documento4 páginasMapa Mental 2Ana JúliaAinda não há avaliações
- Revista Biotec 24Documento44 páginasRevista Biotec 24Claudia Marasciulo100% (1)
- Mobilizaçao NeuralDocumento13 páginasMobilizaçao NeuralTANIAMSMAinda não há avaliações
- Aula 9 - MutaçõesDocumento43 páginasAula 9 - MutaçõesCristina DiasAinda não há avaliações
- Roteiro AAF Sistema NervosoDocumento2 páginasRoteiro AAF Sistema NervosoAdriana BatistaAinda não há avaliações
- Biologia Apontamentos 2Documento22 páginasBiologia Apontamentos 2Ana PaivaAinda não há avaliações
- Aula 11 - Anestesias Superficiais e Infiltrativas - 29-09-2010Documento15 páginasAula 11 - Anestesias Superficiais e Infiltrativas - 29-09-2010Marcos Gomes100% (3)