Você também pode gostar
- Miguel Henrique Freitas FonsecaDocumento17 páginasMiguel Henrique Freitas Fonsecamiguel.lotbrok880Ainda não há avaliações
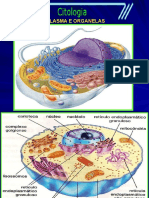
- Organelas CelularesDocumento6 páginasOrganelas CelularesLuara Patrine SalesAinda não há avaliações
- Miguel Henrique Freitas FonsecaDocumento17 páginasMiguel Henrique Freitas Fonsecamiguel.lotbrok880Ainda não há avaliações
- Aula 6 PlastídiosDocumento18 páginasAula 6 Plastídioscalmonhugo24Ainda não há avaliações
- Trabalho Biologia Molecular Constituicao PlastideosDocumento11 páginasTrabalho Biologia Molecular Constituicao PlastideosFlorindoAinda não há avaliações
- A Célula VegetalDocumento2 páginasA Célula VegetalINFORLEXAinda não há avaliações
- Diferenças Entre As Células Animais e VegetaisDocumento1 páginaDiferenças Entre As Células Animais e VegetaisGabi BertiliêAinda não há avaliações
- Célula VegetalDocumento10 páginasCélula VegetalKarollyne Santana PaixãoAinda não há avaliações
- Celula Vegetal1Documento9 páginasCelula Vegetal1CrisSumaqueiroAinda não há avaliações
- Biologia Aula 2Documento8 páginasBiologia Aula 2Pierry IsseiAinda não há avaliações
- Biologia Aula 2Documento8 páginasBiologia Aula 2Kelly DomboAinda não há avaliações
- PESQUISADocumento6 páginasPESQUISAsamara vitoria frias zanellato zanellatoAinda não há avaliações
- Célula EucarionteDocumento4 páginasCélula EucariontesamuelsschAinda não há avaliações
- Pesquisa de BiologiaDocumento4 páginasPesquisa de BiologiaEnoc FerreiraAinda não há avaliações
- Organela Da Célula VegetalDocumento2 páginasOrganela Da Célula VegetalIsabel CamposAinda não há avaliações
- Observação de Grãos de Amido Dentro Da Célula.Documento4 páginasObservação de Grãos de Amido Dentro Da Célula.elizangelacarAinda não há avaliações
- Trabalho de Biologia 12a ClasseDocumento4 páginasTrabalho de Biologia 12a ClasseEduTafulaAinda não há avaliações
- EstruturasDocumento8 páginasEstruturasRofino samuel ernestoAinda não há avaliações
- PlastosDocumento9 páginasPlastosHelena Pastório Dambrós100% (1)
- Citologia Revisado Pontos Importantes Aula de BiologiaDocumento2 páginasCitologia Revisado Pontos Importantes Aula de BiologiaYara FerroAinda não há avaliações
- Ed 1 - Célula VegetalDocumento7 páginasEd 1 - Célula VegetalKassya Lima BrumAinda não há avaliações
- Aula Organelas CursinhoDocumento29 páginasAula Organelas Cursinhobruno.silva.matosAinda não há avaliações
- CITOLOGIA - TaniaDocumento7 páginasCITOLOGIA - TaniageannyyassudaAinda não há avaliações
- Retículo EndoplasmáticoDocumento10 páginasRetículo EndoplasmáticoWillyane Samara Leandro FerreiraAinda não há avaliações
- Cito PlasmaDocumento18 páginasCito PlasmaAndryo CardosoAinda não há avaliações
- Citologia e HistologiaDocumento16 páginasCitologia e HistologiaLuisa AmeduriAinda não há avaliações
- Aula 5Documento13 páginasAula 5Aniceto RosarioAinda não há avaliações
- Resumo Das Ideias Principais Sobre Célula VegetalDocumento3 páginasResumo Das Ideias Principais Sobre Célula VegetalPaulo CezarAinda não há avaliações
- Trabalho de BiologiaDocumento10 páginasTrabalho de BiologiaMaria helena De amorim ferreiraAinda não há avaliações
- OrganelasDocumento3 páginasOrganelasTheus NevesAinda não há avaliações
- Unid 1Documento23 páginasUnid 1renato bryanAinda não há avaliações
- Citologia, Histologia e Embriologia 1Documento23 páginasCitologia, Histologia e Embriologia 1kim TeyanaAinda não há avaliações
- Resumo de Organelas CitoplasmáticasDocumento4 páginasResumo de Organelas CitoplasmáticasAna Laura Campos de PieriAinda não há avaliações
- Biologia - S8Documento6 páginasBiologia - S8Guilherme ColomboAinda não há avaliações
- CélulaDocumento10 páginasCéluladanieleAinda não há avaliações
- Características Gerais Dos Seres VivosDocumento7 páginasCaracterísticas Gerais Dos Seres VivoshericaAinda não há avaliações
- CitologiaDocumento8 páginasCitologiaericc-13hotmail.comAinda não há avaliações
- Aula 3 Citoplasma e OrganelasDocumento37 páginasAula 3 Citoplasma e OrganelassilrogerioAinda não há avaliações
- Organelas CitoplasmáticasDocumento5 páginasOrganelas CitoplasmáticasBrena SousaAinda não há avaliações
- Resumo - GenéticaDocumento16 páginasResumo - Genéticaisabeljulia968Ainda não há avaliações
- CITOPLASMADocumento89 páginasCITOPLASMAVagner Fortes RosadoAinda não há avaliações
- Citologia IiDocumento2 páginasCitologia IiHiroAinda não há avaliações
- Organelas Celulares e Estruturas Celulares.Documento21 páginasOrganelas Celulares e Estruturas Celulares.Daiane LorenzonAinda não há avaliações
- Organelas CelulareDocumento11 páginasOrganelas CelulareGabriel PrataAinda não há avaliações
- Célula VegetalDocumento17 páginasCélula VegetalWanderlei Silva da CunhaAinda não há avaliações
- Citoplasma e Organelas CitoplasmáticasDocumento46 páginasCitoplasma e Organelas Citoplasmáticas0774.rosimeribatistaAinda não há avaliações
- Biologia EnemDocumento29 páginasBiologia EnemJulianna AmorimAinda não há avaliações
- CloropDocumento2 páginasCloroplirio muchidAinda não há avaliações
- Células Procariontes e EucariontesDocumento6 páginasCélulas Procariontes e EucariontesNelson Delú-FilhoAinda não há avaliações
- Aula 01 - IntroduçãoDocumento43 páginasAula 01 - IntroduçãoJordeilson AmaralAinda não há avaliações
- 205949-Aula 2 OrganelasDocumento22 páginas205949-Aula 2 Organelaskeyse rochaAinda não há avaliações
- Aula 7 - Organelas Citoplasmáticas IIDocumento37 páginasAula 7 - Organelas Citoplasmáticas IIhome maiaAinda não há avaliações
- Aula 05 Sistemas de Endomembranas e OrganelasDocumento24 páginasAula 05 Sistemas de Endomembranas e OrganelasBIANCA PIFFERAinda não há avaliações
- Fisiologia VegetalDocumento8 páginasFisiologia VegetalRafael GuardadoAinda não há avaliações
- Citoplasma, HialoplasmaDocumento40 páginasCitoplasma, HialoplasmaJordi GeraldesAinda não há avaliações
- Organelas CitoplasmáticasDocumento5 páginasOrganelas CitoplasmáticasCassia RhayaneAinda não há avaliações
- Aula 1 - Celula VegetalDocumento48 páginasAula 1 - Celula VegetalLaís BarretoAinda não há avaliações
- Diferenças Entre Célula Animal e VegetalDocumento1 páginaDiferenças Entre Célula Animal e VegetalDavi RafaelAinda não há avaliações
- Citologia IDocumento14 páginasCitologia IbjorkAinda não há avaliações
- Estequiometria RevisadoDocumento10 páginasEstequiometria RevisadoMarcelle FelicianoAinda não há avaliações
- Aula 02 Química Geral I ExpDocumento50 páginasAula 02 Química Geral I ExpMarcelle FelicianoAinda não há avaliações
- Óxidos PDFDocumento5 páginasÓxidos PDFMarcelle FelicianoAinda não há avaliações
- Roteiro de Prática 06 - Reações e Propriedades Dos Óxidos - 2017.2 VFDocumento3 páginasRoteiro de Prática 06 - Reações e Propriedades Dos Óxidos - 2017.2 VFMarcelle FelicianoAinda não há avaliações
- Prática Extração Com SoxhletDocumento2 páginasPrática Extração Com SoxhletMarcelle FelicianoAinda não há avaliações
- Tendencias Periodicas Elementos Aula09Documento25 páginasTendencias Periodicas Elementos Aula09Marcelle FelicianoAinda não há avaliações
- Compostos Covalentes - TeoriasDocumento45 páginasCompostos Covalentes - TeoriasMarcelle FelicianoAinda não há avaliações
- Ácidos e BasesDocumento17 páginasÁcidos e BasesMarcelle FelicianoAinda não há avaliações
- Teoria Da Ligacao - de Valencia - Aula 06Documento17 páginasTeoria Da Ligacao - de Valencia - Aula 06Marcelle FelicianoAinda não há avaliações
- Teoria AtômicaDocumento38 páginasTeoria AtômicaMarcelle FelicianoAinda não há avaliações
- Anatomia Região GlúteaDocumento14 páginasAnatomia Região GlúteaAndressa SilvaAinda não há avaliações
- Farmacotécnica BatomDocumento4 páginasFarmacotécnica Batomchagas_83Ainda não há avaliações
- AVC HemorragicoDocumento35 páginasAVC HemorragicoBenigno Tique JonasseAinda não há avaliações
- Crescimento e Renovação CelularDocumento23 páginasCrescimento e Renovação CelularSérgio AmorimAinda não há avaliações
- APH Hemorragias e Choque (MELO e DIAS)Documento42 páginasAPH Hemorragias e Choque (MELO e DIAS)Denis Patricio0% (1)
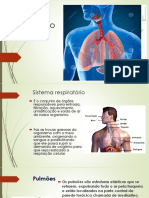
- Sistema RespiratorioDocumento42 páginasSistema RespiratorioThiago Marques Mendes100% (1)
- Bio GeoDocumento9 páginasBio GeoPamela CoxAinda não há avaliações
- Ficha Neuro AdultoDocumento6 páginasFicha Neuro AdultoAntonio Evanir100% (1)
- Aspectos Atuais Sobre Exercício Físico, Fadiga e Nutrição PDFDocumento16 páginasAspectos Atuais Sobre Exercício Físico, Fadiga e Nutrição PDFWander DouglasAinda não há avaliações
- Atividade Laboratorial 1.1Documento3 páginasAtividade Laboratorial 1.1Carla RodriguesAinda não há avaliações
- Fichas Biologia AnoiDocumento102 páginasFichas Biologia AnoiLuis Dias88% (8)
- Biologia Exercicios Fisiologia Animal Sistema Endocrino Gabarito ResolucaoDocumento9 páginasBiologia Exercicios Fisiologia Animal Sistema Endocrino Gabarito ResolucaoMaria Antônia FloresAinda não há avaliações
- Alginac Bula Paciente 21.08.2014Documento25 páginasAlginac Bula Paciente 21.08.2014Leandro MarcosAinda não há avaliações
- ESTUDO de CASO (Processos Biológicos)Documento11 páginasESTUDO de CASO (Processos Biológicos)nina33250Ainda não há avaliações
- UFCD - 7211 - Os Sistemas Do Corpo Humano - Imunitário, Circulatório, Respiratório, Nervoso e Músculo-Esquelético - ÍndiceDocumento5 páginasUFCD - 7211 - Os Sistemas Do Corpo Humano - Imunitário, Circulatório, Respiratório, Nervoso e Músculo-Esquelético - ÍndiceMANUAIS FORMAÇÃOAinda não há avaliações
- Ejercicios de OrganicaDocumento11 páginasEjercicios de OrganicaLuuLi PeréAinda não há avaliações
- Parasitarias 2Documento15 páginasParasitarias 2Luiz FelipeAinda não há avaliações
- Recuperação AvaliaçãoDocumento3 páginasRecuperação AvaliaçãoDaniel FerreiraAinda não há avaliações
- Slides Trabalho Teoria CelularDocumento33 páginasSlides Trabalho Teoria CelularBrunoLopesAinda não há avaliações
- Efeitos Do Tribulus Terrestris PDFDocumento164 páginasEfeitos Do Tribulus Terrestris PDFAnderson F MartinezAinda não há avaliações
- 8 Aula - GoniometriaDocumento37 páginas8 Aula - GoniometriaLeonardo LealAinda não há avaliações
- Sistema MuscularDocumento5 páginasSistema Muscularelaine santosAinda não há avaliações
- BrassicasDocumento6 páginasBrassicasAnna Carolina DominguesAinda não há avaliações
- Protocolo - Contagem de Eritrócitos e Contagem Total de LeucócitosDocumento3 páginasProtocolo - Contagem de Eritrócitos e Contagem Total de LeucócitosMariana NóbregaAinda não há avaliações
- Diagnóstico e Datação Da GravidezDocumento3 páginasDiagnóstico e Datação Da GravidezTiago SuaresAinda não há avaliações
- Farmacologia Dos OpióidesDocumento15 páginasFarmacologia Dos OpióidesAmanda Hoffmann100% (1)
- Plano de Ensino Fisiologia 2Documento5 páginasPlano de Ensino Fisiologia 2LaísAinda não há avaliações
- Ivan Pavlov BiografiaDocumento2 páginasIvan Pavlov BiografiaMauricio Daniel Colete100% (1)
- Sistema Muscular - Parte 1: 6 ANO Aula 14 - 3 BimestreDocumento24 páginasSistema Muscular - Parte 1: 6 ANO Aula 14 - 3 Bimestreemillynascimentos508Ainda não há avaliações
- Pulsion - 28-06-23Documento70 páginasPulsion - 28-06-23João Gabriel AlencarAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Dieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNo EverandDieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNota: 5 de 5 estrelas5/5 (4)
- Uma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNo EverandUma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNota: 4.5 de 5 estrelas4.5/5 (9)
- Hackeando Darwin: Engenharia genética e o futuro da humanidadeNo EverandHackeando Darwin: Engenharia genética e o futuro da humanidadeAinda não há avaliações
- O Guia Da Apicultura Para Iniciantes: O Guia Completo Para Criar Abelhas, Aumentar Suas Colônias E Fazer Sua Colmeia ProsperarNo EverandO Guia Da Apicultura Para Iniciantes: O Guia Completo Para Criar Abelhas, Aumentar Suas Colônias E Fazer Sua Colmeia ProsperarAinda não há avaliações
- Fundamentos Em Cirurgia OralNo EverandFundamentos Em Cirurgia OralNota: 5 de 5 estrelas5/5 (1)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Fitoenergética: A Energia das Plantas no Equilíbrio da AlmaNo EverandFitoenergética: A Energia das Plantas no Equilíbrio da AlmaNota: 5 de 5 estrelas5/5 (17)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações