Você também pode gostar
- Transferencia UfprDocumento6 páginasTransferencia UfprRicardo Gomes Fernandes100% (1)
- Raças e Linhagens de AvesDocumento52 páginasRaças e Linhagens de AvesJ SilvestriAinda não há avaliações
- Resumo AV1 e AV2 - Embriologia e HistologiaDocumento11 páginasResumo AV1 e AV2 - Embriologia e HistologiaLuciene BarretoAinda não há avaliações
- Krishnamurti, J. - Que o Entendimento Seja LeiDocumento7 páginasKrishnamurti, J. - Que o Entendimento Seja LeiUpasakaAinda não há avaliações
- Pires - Tutoria Reprodução P1Documento31 páginasPires - Tutoria Reprodução P1pauloAinda não há avaliações
- Líquidos Intra e ExtracelularesDocumento31 páginasLíquidos Intra e ExtracelularesDamom Ribeiro100% (2)
- Defesa MultaDocumento4 páginasDefesa MultaRicardo Gomes FernandesAinda não há avaliações
- Processo de Fabricação Do CouroDocumento9 páginasProcesso de Fabricação Do CouroJuliana RodriguesAinda não há avaliações
- Anatomia Dos Rins PDFDocumento22 páginasAnatomia Dos Rins PDFWrdyley VasconcelosAinda não há avaliações
- Lista de Livros Censurados PDFDocumento9 páginasLista de Livros Censurados PDFPsycho_AntichristAinda não há avaliações
- Lesões Reversíveis e IrreversíveisDocumento67 páginasLesões Reversíveis e IrreversíveisjaquelinessAinda não há avaliações
- Livros - Livros em Português - Versão 8Documento11 páginasLivros - Livros em Português - Versão 8ACP F100% (1)
- Aula 5 - 4 A 8 Semana de Desenvolvimento EmbrionárioDocumento52 páginasAula 5 - 4 A 8 Semana de Desenvolvimento EmbrionárioDavid MendesAinda não há avaliações
- A Bíblia e A SaúdeDocumento2 páginasA Bíblia e A SaúdesivanildoAinda não há avaliações
- Reação Antígeno-Anticorpo e o Complemento em ImunohematologiaDocumento23 páginasReação Antígeno-Anticorpo e o Complemento em ImunohematologiaJosé Alisson dos Santos100% (2)
- Resumo Endocrino GuytonDocumento18 páginasResumo Endocrino GuytonWaldman Santos100% (1)
- 1-Imuno-Hematologia 2009Documento88 páginas1-Imuno-Hematologia 2009José Alisson dos SantosAinda não há avaliações
- Um Belo Hadith Do Profeta Muhammad S.A.A.SDocumento5 páginasUm Belo Hadith Do Profeta Muhammad S.A.A.Sلا اله الا الله محمد رسول الله 1100% (1)
- Hemocitopoese - ResumoDocumento7 páginasHemocitopoese - ResumoRichardson Oliveira50% (2)
- Aula Membrana Plasmatica Aula 3Documento46 páginasAula Membrana Plasmatica Aula 3quel04Ainda não há avaliações
- O Valor Da Vida (Aborto)Documento22 páginasO Valor Da Vida (Aborto)Allan MateusAinda não há avaliações
- Divisões CelularesDocumento22 páginasDivisões CelularesFabio Gonçalves Da SilvaAinda não há avaliações
- Alcorao o Milagre Linguistico - Sheikh Ahmad MazloumDocumento54 páginasAlcorao o Milagre Linguistico - Sheikh Ahmad MazloumDany Aziz100% (1)
- 10-Aula Histoligia AV2Documento26 páginas10-Aula Histoligia AV2Gilberto ConstantinoAinda não há avaliações
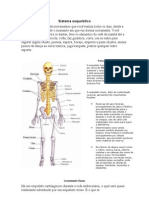
- Sistema EsqueléticoDocumento11 páginasSistema EsqueléticoAgnariane S Moreira100% (1)
- Sistema Nervoso SimpáticoDocumento11 páginasSistema Nervoso SimpáticoMateus Dos Santos BandeiraAinda não há avaliações
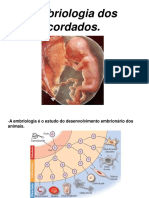
- Embriologia Dos CordadosDocumento11 páginasEmbriologia Dos CordadosRicardo Carol LoboAinda não há avaliações
- Embriologia Da Quarta A Oitava SemanasDocumento29 páginasEmbriologia Da Quarta A Oitava SemanasAlexson PaixaoAinda não há avaliações
- TireóideDocumento12 páginasTireóideGabriel Felipe Das NevesAinda não há avaliações
- Agenda Pós-Neoliberal - Todas As EdiçõesDocumento369 páginasAgenda Pós-Neoliberal - Todas As EdiçõesIbase Na RedeAinda não há avaliações
- Microbiologia - Fundamentos e Perspectivas 4 EdDocumento738 páginasMicrobiologia - Fundamentos e Perspectivas 4 Edgéssica100% (2)
- PQQ - A Origem Da VidaDocumento8 páginasPQQ - A Origem Da VidaJoao Luiz Darques FerreiraAinda não há avaliações
- Funções Secretoras Do Trato AlimentarDocumento10 páginasFunções Secretoras Do Trato AlimentarLeticia FariasAinda não há avaliações
- Membrana PlasmáticaDocumento84 páginasMembrana Plasmáticaphaela_maAinda não há avaliações
- Hormonio de CrescimentoDocumento41 páginasHormonio de CrescimentoRobinsom Alves100% (1)
- Sistema EsqueléticoDocumento3 páginasSistema EsqueléticoKanasa 03Ainda não há avaliações
- Resumo - Hematopoese e Eritropoese - HematologiaDocumento14 páginasResumo - Hematopoese e Eritropoese - Hematologiadanimb100% (1)
- HomeostaseDocumento24 páginasHomeostaseYanesAinda não há avaliações
- Resumo FisiologiaDocumento23 páginasResumo FisiologiaGabrielAinda não há avaliações
- Extensivoenem Biologia2 Mutações Cromossômicas 10-06-2019Documento6 páginasExtensivoenem Biologia2 Mutações Cromossômicas 10-06-2019Pedro ManoelAinda não há avaliações
- Citologia HormonalDocumento4 páginasCitologia HormonalLeonardo Daniel MendesAinda não há avaliações
- Atividade Contextualizada (AOL5) - BIOFISICADocumento1 páginaAtividade Contextualizada (AOL5) - BIOFISICAJessica Teodoro da MotaAinda não há avaliações
- Capitulo 32 GuytonDocumento7 páginasCapitulo 32 GuytonRenan Helmer Noelves0% (1)
- Estudo SanguíneoDocumento12 páginasEstudo SanguíneoLucas De Godoy BuenoAinda não há avaliações
- Questionário Sobre RenalDocumento20 páginasQuestionário Sobre RenalAlmir Fernandes de Carvalho100% (8)
- Placenta MedicinaDocumento49 páginasPlacenta MedicinaBruno Avelar100% (1)
- Endocitose e ExocitoseDocumento2 páginasEndocitose e ExocitoseJoana DâmasoAinda não há avaliações
- Atlas de UroDocumento18 páginasAtlas de UroFernanda ZanellaAinda não há avaliações
- Arq Nac OVNIS Listagem ARXDocumento248 páginasArq Nac OVNIS Listagem ARXलुकास विडालAinda não há avaliações
- Sistema Reprodutor MasculinoDocumento11 páginasSistema Reprodutor MasculinoAssistência Social ingazeiraAinda não há avaliações
- Resumo FarmacologiaDocumento17 páginasResumo FarmacologiaJoão Vitor Fernandes da CunhaAinda não há avaliações
- Fisiologia Comparada - Circulação S.19Documento15 páginasFisiologia Comparada - Circulação S.19Rosana Galev OliveiraAinda não há avaliações
- Fisiologia - Fisiologia Do SangueDocumento68 páginasFisiologia - Fisiologia Do SangueDeborah Diniz100% (2)
- MicrocirculaçãoDocumento31 páginasMicrocirculaçãoReinaldo RiquetAinda não há avaliações
- Proteínas Da Membrana EritrocitáriaDocumento36 páginasProteínas Da Membrana EritrocitáriaBruno Felipe100% (2)
- Revisao Tecido Muscular e NervosoDocumento14 páginasRevisao Tecido Muscular e NervosoAndrezza CristinaAinda não há avaliações
- Teste de Conhecimentos em Genética ForenseDocumento13 páginasTeste de Conhecimentos em Genética ForenseNatália Marcondes100% (1)
- Resumo Hematologia BásicaDocumento9 páginasResumo Hematologia BásicaBiouma MadeiraAinda não há avaliações
- Relatório Músculo LisoDocumento9 páginasRelatório Músculo LisoSilas PintoAinda não há avaliações
- Compilação Resumos Fisiologia UrinárioDocumento31 páginasCompilação Resumos Fisiologia UrinárioFernandoRibeiroAinda não há avaliações
- AULA DE BIOELETROGÊNESE 2014 AlunosDocumento53 páginasAULA DE BIOELETROGÊNESE 2014 AlunosLhana Lorena Melo AtanasioAinda não há avaliações
- CitologiaDocumento10 páginasCitologiaronaldotyskiAinda não há avaliações
- Requerimento de Alteração de EndereçoDocumento1 páginaRequerimento de Alteração de EndereçoRicardo Gomes FernandesAinda não há avaliações
- Questões Objetivas Independencia BrasilDocumento4 páginasQuestões Objetivas Independencia BrasilRicardo Gomes FernandesAinda não há avaliações
- Agamben. O AbertoDocumento14 páginasAgamben. O AbertoCaio Souto100% (1)
- Lquidos Serosos PDFDocumento17 páginasLquidos Serosos PDFjorgedasaude6025100% (1)
- 3º Período Teste 6º AnoDocumento2 páginas3º Período Teste 6º Anoarlindoarantes4845Ainda não há avaliações
- Apostila Zootecnia Geral 2010 PDFDocumento60 páginasApostila Zootecnia Geral 2010 PDFJosé Cutrim Junior100% (2)
- HelmintologiaDocumento7 páginasHelmintologiaThayna De AssisAinda não há avaliações
- Diagnóstico Por Imagem Do Pâncreas PDFDocumento29 páginasDiagnóstico Por Imagem Do Pâncreas PDFPaulo Henrique Haß FilhoAinda não há avaliações
- O Cultivo de Ervas MedicinaisDocumento8 páginasO Cultivo de Ervas MedicinaisMircia RomanAinda não há avaliações
- cONCIENCIA fONOLOGICADocumento17 páginascONCIENCIA fONOLOGICADaiMenezesAinda não há avaliações
- Atividades FevereiroDocumento14 páginasAtividades FevereiroDaniela Fernandes Pimenta dos AnjosAinda não há avaliações
- pt7 PPT 13 RelacoesDocumento11 páginaspt7 PPT 13 RelacoesProf.ª Maria de Fatima Martins de VasconcelosAinda não há avaliações
- Texto de Leitura e ExerciciosDocumento2 páginasTexto de Leitura e ExerciciosFlavia MacedoAinda não há avaliações
- Fisiologia e Metabolismo de Sistema OsseoDocumento19 páginasFisiologia e Metabolismo de Sistema OsseoAnderson De Oliveira SilvaAinda não há avaliações
- Hemoglobin ADocumento20 páginasHemoglobin Aalice duarteAinda não há avaliações
- Atividade 1º e 2º Ano Libras StellaDocumento74 páginasAtividade 1º e 2º Ano Libras StellaCarlos Pereira da SilvaAinda não há avaliações
- Vigilância, Prevenção E Controle Das Zoonoses: Maiara Leite BarberinoDocumento21 páginasVigilância, Prevenção E Controle Das Zoonoses: Maiara Leite BarberinoAndréa CostaAinda não há avaliações
- Aula 9 - Ascaris LumbricoidesDocumento31 páginasAula 9 - Ascaris LumbricoidesAlda Almeida100% (1)
- Atividades Da Semana 1 AlteraçãoDocumento6 páginasAtividades Da Semana 1 AlteraçãoSolange MarizAinda não há avaliações
- Sistema Sensorial RelatorioDocumento23 páginasSistema Sensorial RelatorioAdrea Carvalho Oliveira100% (1)
- Eu Quero Desenhar Do ZeroDocumento73 páginasEu Quero Desenhar Do Zeroftmoraes100% (3)
- Anatomia 7Documento29 páginasAnatomia 7Anna Carolina MacedoAinda não há avaliações
- DiversasDocumento32 páginasDiversasSilvanaAinda não há avaliações
- Planilha Custos Gran FatiêDocumento9 páginasPlanilha Custos Gran FatiêdouglasacahuAinda não há avaliações
- Ecologia 1Documento2 páginasEcologia 1teatica6754Ainda não há avaliações
- Apostila Paleontologia Bsica 2 Prova PrCambriano Ao FanerozicoDocumento25 páginasApostila Paleontologia Bsica 2 Prova PrCambriano Ao FanerozicoIsabelle VasconcelosAinda não há avaliações
- 6º Ano EF Português Júlia PDFDocumento33 páginas6º Ano EF Português Júlia PDFJéssica OliveiraAinda não há avaliações
- Ficha Do Criador Editavel 1Documento1 páginaFicha Do Criador Editavel 1Pedro AssisAinda não há avaliações
- 3o Simulado PMMSCursoMazziotti-18Documento13 páginas3o Simulado PMMSCursoMazziotti-18Karoline GimenezAinda não há avaliações
- Atividade para Aula de 5º Ano Do Ensino FundamentalDocumento3 páginasAtividade para Aula de 5º Ano Do Ensino FundamentalMarcia OdaraAinda não há avaliações