Você também pode gostar
- Práticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNo EverandPráticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNota: 4 de 5 estrelas4/5 (1)
- Degeneração TesticularDocumento7 páginasDegeneração TesticulargcguilhermAinda não há avaliações
- Retinol na Produção In Vitro de Embriões CaprinosNo EverandRetinol na Produção In Vitro de Embriões CaprinosAinda não há avaliações
- Patologias EspermáticasDocumento11 páginasPatologias EspermáticasMichelle NaomiAinda não há avaliações
- HistologiaPL - Testículo, Espermatozóide e Qualidade EspermáticaDocumento23 páginasHistologiaPL - Testículo, Espermatozóide e Qualidade Espermáticaviolleta gomiaAinda não há avaliações
- Espermograma e Espermocultura Na BovinoculturaDocumento5 páginasEspermograma e Espermocultura Na Bovinoculturaadrianwelinton458Ainda não há avaliações
- Componentes Do Plasma Seminal e Sua Influc3aancia Sobre A Criopreservac3a7c3a3o e Fertilidade de Espermatozc3b3ides Equinos1Documento12 páginasComponentes Do Plasma Seminal e Sua Influc3aancia Sobre A Criopreservac3a7c3a3o e Fertilidade de Espermatozc3b3ides Equinos1Higino ValentimAinda não há avaliações
- 1 Desen Embrio e Difer SexualDocumento17 páginas1 Desen Embrio e Difer SexualChácara Arte SuaveAinda não há avaliações
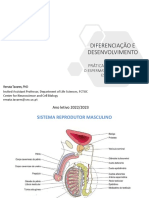
- Sistema Reprodutor Masculino e GametogêneseDocumento2 páginasSistema Reprodutor Masculino e GametogêneseDanielle MariaAinda não há avaliações
- Trabalho Escrito Fisiopatologia Da ReproduçãoDocumento19 páginasTrabalho Escrito Fisiopatologia Da ReproduçãoPablo CavalcantiAinda não há avaliações
- Aspectos Relevantes Na Avaliação Da Morfologia Espermática Do SuínoDocumento3 páginasAspectos Relevantes Na Avaliação Da Morfologia Espermática Do SuínoElisa da CunhaAinda não há avaliações
- 10 - Embriologia HumanaDocumento191 páginas10 - Embriologia HumanaRafael RodriguesAinda não há avaliações
- Freemartinismo em BovinosDocumento65 páginasFreemartinismo em BovinosEliziana E. S.Ainda não há avaliações
- Freemartinismo em Bovinos: Revisão de Literatura Freemartins in Cattle: A ReviewDocumento9 páginasFreemartinismo em Bovinos: Revisão de Literatura Freemartins in Cattle: A ReviewJapah MeloAinda não há avaliações
- EspermatogêneseDocumento52 páginasEspermatogêneseGabrielleGesser100% (1)
- Histologia e Embriologia - Aula 2Documento15 páginasHistologia e Embriologia - Aula 2cunhadarvinAinda não há avaliações
- Trabalho de Conclusão de Curso: Centro Universitário Campo Limpo PaulistaDocumento18 páginasTrabalho de Conclusão de Curso: Centro Universitário Campo Limpo PaulistaRafael BorgesAinda não há avaliações
- Andrologia e Criopreserção de Semem BovinoDocumento8 páginasAndrologia e Criopreserção de Semem BovinoThalita Masoti BlankenheimAinda não há avaliações
- Embriologia A1Documento2 páginasEmbriologia A1luana_cfpAinda não há avaliações
- Ae bg11 Teste Avaliacao 2 Nov 2020Documento10 páginasAe bg11 Teste Avaliacao 2 Nov 2020RicardoAinda não há avaliações
- GametogêneseDocumento4 páginasGametogêneseLays MonteiroAinda não há avaliações
- Teste Avaliacao 2 Nov 2020Documento10 páginasTeste Avaliacao 2 Nov 2020antonia44322100% (2)
- 000Documento7 páginas000stepoutnowAinda não há avaliações
- Ovário Policístico PDFDocumento2 páginasOvário Policístico PDFemanuelAinda não há avaliações
- 1.aparelho Reprodutor MasculinoDocumento23 páginas1.aparelho Reprodutor MasculinoAna Bz CruzAinda não há avaliações
- Anat.e Fisiol - Sistema Reprodutor Masculino 19pgDocumento19 páginasAnat.e Fisiol - Sistema Reprodutor Masculino 19pgderek0% (1)
- Ud I - Sexualidade Humana PDFDocumento80 páginasUd I - Sexualidade Humana PDFPedro HenriqueAinda não há avaliações
- Infertilidade MasculinaDocumento15 páginasInfertilidade Masculinacatarina rodriguesAinda não há avaliações
- Esta GioDocumento22 páginasEsta GioGeovana AndradeAinda não há avaliações
- Aula 1 - Características Anatômicas e Citológicas Do Trato Genital Feminino - Citologia Clínica - FMNDocumento57 páginasAula 1 - Características Anatômicas e Citológicas Do Trato Genital Feminino - Citologia Clínica - FMNAlexsandra CruzAinda não há avaliações
- Espermatozoides PDFDocumento9 páginasEspermatozoides PDFjanjardins23Ainda não há avaliações
- Santos - 2012 - Efeito Do Tipo de Fixador e Tempo de Fixação Na Morfologia de Folículos Ovarianos Pré-AntraisDocumento8 páginasSantos - 2012 - Efeito Do Tipo de Fixador e Tempo de Fixação Na Morfologia de Folículos Ovarianos Pré-Antraisevelynandrade25Ainda não há avaliações
- Apostila Andrologico Biotran - PRNDocumento27 páginasApostila Andrologico Biotran - PRNHenry Chinaski100% (1)
- Terato G Én Ios 2023Documento28 páginasTerato G Én Ios 2023gffgAinda não há avaliações
- PDF Imprimir 2 MarcadoresDocumento14 páginasPDF Imprimir 2 Marcadoresenzodatrindade2512Ainda não há avaliações
- Gametogênese HumanaDocumento8 páginasGametogênese HumanaRogerioGuimaraesAinda não há avaliações
- Gametogênese e FertilizaçãoDocumento38 páginasGametogênese e FertilizaçãoLarissa AlvesAinda não há avaliações
- Potencial Reprodutivo em Tamanduá-Bandeira - Maíra C FerreiraDocumento19 páginasPotencial Reprodutivo em Tamanduá-Bandeira - Maíra C Ferreiramairacf190400Ainda não há avaliações
- Infertilidade MasculinaDocumento3 páginasInfertilidade Masculinacatarina rodriguesAinda não há avaliações
- 1 Série: Ciências Da Natureza (Biologia, Química E Física)Documento7 páginas1 Série: Ciências Da Natureza (Biologia, Química E Física)Thaylla KarinAinda não há avaliações
- Roteiro de Estudo EmbriologiaDocumento8 páginasRoteiro de Estudo EmbriologiaMarcos Marzano100% (1)
- Teste de Avaliacao Escrita de Biologia 12 1 ReservaDocumento7 páginasTeste de Avaliacao Escrita de Biologia 12 1 ReservaRenato Costa100% (21)
- EmbriologiaDocumento44 páginasEmbriologiaJúlia CarolinaAinda não há avaliações
- Embriologia 2011 - GabaritoDocumento2 páginasEmbriologia 2011 - GabaritoIvaneaVasquesCruzAinda não há avaliações
- 1 gametogeneseII2008Documento24 páginas1 gametogeneseII2008Aldo Brásio SimãoAinda não há avaliações
- Infertilidade Masculina 2Documento6 páginasInfertilidade Masculina 2spindolaAinda não há avaliações
- Biologia Celular - Formação Do EspermaDocumento8 páginasBiologia Celular - Formação Do EspermaKolynAinda não há avaliações
- Roteiros de Aula PraticaDocumento24 páginasRoteiros de Aula PraticaEliane CardosoAinda não há avaliações
- Roteiro de Estudo Mód. I - Crescimento e Desenvolvimento IDocumento10 páginasRoteiro de Estudo Mód. I - Crescimento e Desenvolvimento IAlanys MattosAinda não há avaliações
- Questionario 9Documento4 páginasQuestionario 9Bruna SoaresAinda não há avaliações
- Oogenese e Modificações EpigenéticasDocumento8 páginasOogenese e Modificações EpigenéticasMariane Gabriela Cesar Ribeiro FerreiraAinda não há avaliações
- Introdução À EmbriologiaDocumento38 páginasIntrodução À EmbriologiaSusana AlvesAinda não há avaliações
- Biologia Final 12ºDocumento28 páginasBiologia Final 12ºbeatrizlorenadossantosAinda não há avaliações
- Aula 1 - Segmentação e BlastulaDocumento50 páginasAula 1 - Segmentação e BlastulalahfmchAinda não há avaliações
- Como A Morfologia Dos Espermatozoides AfetaDocumento5 páginasComo A Morfologia Dos Espermatozoides AfetaLaura SantosAinda não há avaliações
- Gomes Et Al., 2015. Exame Andrologico em BovinosDocumento6 páginasGomes Et Al., 2015. Exame Andrologico em Bovinosrodrigoferrazza1662Ainda não há avaliações
- Trabalho GenéticaDocumento6 páginasTrabalho GenéticaLuciana Dalla ChiesaAinda não há avaliações
- EmbriologiaDocumento4 páginasEmbriologiaDanielle MariaAinda não há avaliações
- Circular Tecnica Embrapa - A Importancia Do Exame Andrologico em Bovinos PDFDocumento13 páginasCircular Tecnica Embrapa - A Importancia Do Exame Andrologico em Bovinos PDFrodrigoferrazza1662Ainda não há avaliações
- Diabetes MellitusDocumento21 páginasDiabetes MellitusLuana milena AmaranteAinda não há avaliações
- Apostila Curso Biotecnologias ReprodutivasDocumento21 páginasApostila Curso Biotecnologias Reprodutivasrodrigoferrazza1662Ainda não há avaliações
- Cap 7 FisiocicloestralDocumento31 páginasCap 7 FisiocicloestralBeatriz DuarteAinda não há avaliações
- Rtoniolli,+09 +18 +REVISÃO+-+2021+-+pg+85+a+100Documento16 páginasRtoniolli,+09 +18 +REVISÃO+-+2021+-+pg+85+a+100Luana milena AmaranteAinda não há avaliações
- Gineco ObstetriciaDocumento69 páginasGineco ObstetriciaLuiza Maria BeckenkampAinda não há avaliações
- Mae MenininhaDocumento14 páginasMae MenininhaOdonel SerranoAinda não há avaliações
- 182 544 1 PB - 221021 - 083725Documento12 páginas182 544 1 PB - 221021 - 083725Laura Prado CarneiroAinda não há avaliações
- Sangramentos Da Primeira Metade Da GestaçãoDocumento56 páginasSangramentos Da Primeira Metade Da GestaçãoDanni MenezesAinda não há avaliações
- Orientações Do Sistema Alojamento Conjunto - 2018Documento4 páginasOrientações Do Sistema Alojamento Conjunto - 2018Jean Carlos silva pinheiroAinda não há avaliações
- Aparelho Reprodutor FemininoDocumento34 páginasAparelho Reprodutor FemininoAna Beatriz AlmeidaAinda não há avaliações
- EVARISTO, Conceição. Insubmissas Lágrimas de MulheresDocumento58 páginasEVARISTO, Conceição. Insubmissas Lágrimas de MulheresMarcosMeinerzAinda não há avaliações
- Gineco 5 - Sangramento Uterino AnormalDocumento10 páginasGineco 5 - Sangramento Uterino AnormalpauloAinda não há avaliações
- Hormônios Sexuais e Métodos ContraceptivosDocumento13 páginasHormônios Sexuais e Métodos ContraceptivosDêh Andrade100% (1)
- Comportamento SuinoDocumento27 páginasComportamento SuinoNeoro RodriguesAinda não há avaliações
- A Doença Inflamatória Pélvica (DIP) : Fatores de RiscoDocumento8 páginasA Doença Inflamatória Pélvica (DIP) : Fatores de RiscoMaria Gabriella Andrade BonaAinda não há avaliações
- Uma Vovó de Outro MundoDocumento5 páginasUma Vovó de Outro MundoRebecca BorgesAinda não há avaliações
- Atividade Pratica Ciencias-Morfofuncionais.Documento16 páginasAtividade Pratica Ciencias-Morfofuncionais.Fransley PereiraAinda não há avaliações
- Laqueadura e Vasectomia - Laudo e ParecerDocumento3 páginasLaqueadura e Vasectomia - Laudo e ParecerSidnei SilvaAinda não há avaliações
- Mesa Real Segundo Odete Lopes MazzaDocumento7 páginasMesa Real Segundo Odete Lopes MazzaJacqueline Souza100% (1)
- 8o ANO CIENCIAS 1Documento2 páginas8o ANO CIENCIAS 1Leanderson CalixtoAinda não há avaliações
- Anatomia e Biomecânica Do Assoalho PélvicoDocumento49 páginasAnatomia e Biomecânica Do Assoalho Pélvicojack100% (1)
- Resumo - Sistema Reprodutor MasculinoDocumento3 páginasResumo - Sistema Reprodutor MasculinoPedro DuarteAinda não há avaliações
- Mapa Mental - Desenvolvimento EmbrionárioDocumento5 páginasMapa Mental - Desenvolvimento EmbrionárioVanessa MedeirosAinda não há avaliações
- Paternidade e Medicina Forensx03042012152105Documento20 páginasPaternidade e Medicina Forensx03042012152105Andressa SilvaAinda não há avaliações
- Anatomia GestaçãoDocumento19 páginasAnatomia GestaçãoEnzo CobucciAinda não há avaliações
- Classificaçao Do RNDocumento6 páginasClassificaçao Do RN666Ozzy666Ainda não há avaliações
- Bio 030 Vpmednasuacasa - ExercíciosDocumento3 páginasBio 030 Vpmednasuacasa - ExercíciosNatalia RibeiroAinda não há avaliações
- Hemograma AutomatizadoDocumento4 páginasHemograma AutomatizadojjoelmaguerraAinda não há avaliações
- Produção de CoelhosDocumento31 páginasProdução de CoelhosRafaela FonsecaAinda não há avaliações
- Métodos ContraceptivosDocumento1 páginaMétodos ContraceptivosYASMIM DINIZAinda não há avaliações
- Estimativa Atualizada - Fiv.102022Documento1 páginaEstimativa Atualizada - Fiv.102022Mirian Camanes VidalAinda não há avaliações
- Livro de Posições Do ShiiuDocumento29 páginasLivro de Posições Do ShiiuCherry Jett BombAinda não há avaliações
- 2c2bateste 12a 2011-2012Documento4 páginas2c2bateste 12a 2011-2012Maria IsabelAinda não há avaliações
- Ciclo Menstrual - MindMeister Mapa MentalDocumento1 páginaCiclo Menstrual - MindMeister Mapa MentalBruno MauricioAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Hackeando Darwin: Engenharia genética e o futuro da humanidadeNo EverandHackeando Darwin: Engenharia genética e o futuro da humanidadeAinda não há avaliações
- Aprender mais rápido: Habilidades de estudo, memória e neurociênciaNo EverandAprender mais rápido: Habilidades de estudo, memória e neurociênciaNota: 4 de 5 estrelas4/5 (5)
- Uma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNo EverandUma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNota: 4.5 de 5 estrelas4.5/5 (9)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- O Licenciamento Ambiental no Brasil na PráticaNo EverandO Licenciamento Ambiental no Brasil na PráticaNota: 1 de 5 estrelas1/5 (2)
- Dieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNo EverandDieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNota: 5 de 5 estrelas5/5 (4)
- Tudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNo EverandTudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNota: 4 de 5 estrelas4/5 (5)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)