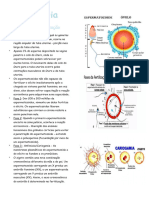
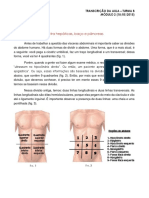
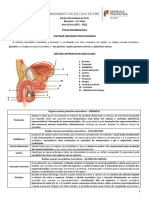
Você também pode gostar
- Primeiras Semanas EMBRIODocumento3 páginasPrimeiras Semanas EMBRIOColégio FochAinda não há avaliações
- (EMBRIOLOGIA - RESUMO) GastrulaçãoDocumento12 páginas(EMBRIOLOGIA - RESUMO) GastrulaçãoCaroline MilheiroAinda não há avaliações
- Desenvolvimento embrionário e formação das camadas germinativasDocumento43 páginasDesenvolvimento embrionário e formação das camadas germinativaslaercio matheusAinda não há avaliações
- Zoologia GeralDocumento13 páginasZoologia GeralGo Cuambe OrlandoAinda não há avaliações
- DESENVOLVIMENTO HUMANODocumento6 páginasDESENVOLVIMENTO HUMANOPaulo RodrigoAinda não há avaliações
- Introdução À Histologia e Embriologia HumanaDocumento4 páginasIntrodução À Histologia e Embriologia HumanaLeticia António AntónioAinda não há avaliações
- Resumo 1 A 8 Semana Do DesenvolvimentoDocumento7 páginasResumo 1 A 8 Semana Do DesenvolvimentoLuiza LeseAinda não há avaliações
- ECTO MESO E ENDO Folheto Embrionário PDFDocumento8 páginasECTO MESO E ENDO Folheto Embrionário PDFfassisbr6137Ainda não há avaliações
- Embriologia GeralDocumento13 páginasEmbriologia GeralabresserAinda não há avaliações
- Ebook - Extensivo Medicina - Semana 43 CompactadoDocumento294 páginasEbook - Extensivo Medicina - Semana 43 CompactadodjeifnaAinda não há avaliações
- ED Biomorfo PR2 GabaritoDocumento5 páginasED Biomorfo PR2 Gabaritomaria vitoriaAinda não há avaliações
- Desenvolvimento EmbrionárioDocumento6 páginasDesenvolvimento EmbrionárioRian silvaAinda não há avaliações
- Desenvolvimento fetal na primeira e segunda semanaDocumento19 páginasDesenvolvimento fetal na primeira e segunda semanaRafael de Lima SantosAinda não há avaliações
- W - HistólogiaDocumento10 páginasW - HistólogiaPedro ThaweAinda não há avaliações
- Principais Eventos Da Segunda e Terceira Semana Do Desenvolvimento EmbrionárioDocumento6 páginasPrincipais Eventos Da Segunda e Terceira Semana Do Desenvolvimento EmbrionárioPaolaRaeski100% (1)
- EmbriologiaDocumento7 páginasEmbriologiamila.rgamaAinda não há avaliações
- EMBRIOLOGIADocumento16 páginasEMBRIOLOGIAFernanda Rocha DinizAinda não há avaliações
- Desenvolvimento embrionário da 3a à 8a semanaDocumento10 páginasDesenvolvimento embrionário da 3a à 8a semanaEmanuel NatanaelAinda não há avaliações
- ED - Fisio Embrio p2Documento4 páginasED - Fisio Embrio p2Felipe RomãoAinda não há avaliações
- Embriologia HumanaDocumento5 páginasEmbriologia HumanaEricka SantosAinda não há avaliações
- Aula 3. Gastrulacao - 3 SemanaDocumento43 páginasAula 3. Gastrulacao - 3 SemanaAuryelle Sousa AzevedoAinda não há avaliações
- Diferenciação Dos Folhetos Embrionarios - SanarDocumento9 páginasDiferenciação Dos Folhetos Embrionarios - SanarMel na MeddAinda não há avaliações
- Histologia e EmbriologiaDocumento16 páginasHistologia e Embriologialulu.helena2806Ainda não há avaliações
- Arquitetura animal e origem do mesodermaDocumento19 páginasArquitetura animal e origem do mesodermaUelinton SantanaAinda não há avaliações
- A Ressignificação para o Gêmeo SobreviventeDocumento47 páginasA Ressignificação para o Gêmeo SobreviventeCrislainny RodriguesAinda não há avaliações
- Desenvolvimento Embrionário 1-3 SemDocumento5 páginasDesenvolvimento Embrionário 1-3 SemleticiaAinda não há avaliações
- Embriologia Humana - Revisão de Gametogênese e Desenvolvimento EmbrionárioDocumento3 páginasEmbriologia Humana - Revisão de Gametogênese e Desenvolvimento EmbrionárioMalu MedAinda não há avaliações
- Processo de Fecundação emDocumento53 páginasProcesso de Fecundação emines capelaAinda não há avaliações
- A Gastrulação É Um Estágio Importante Do Desenvolvimento EmbrionárioDocumento3 páginasA Gastrulação É Um Estágio Importante Do Desenvolvimento EmbrionárioAugusto UggioniAinda não há avaliações
- Questões de embriologia humanaDocumento3 páginasQuestões de embriologia humanaMilla Rodrigues40% (5)
- Embriologia GastrulaçãoDocumento49 páginasEmbriologia GastrulaçãoLucélia BatalhaAinda não há avaliações
- Descrevendo fertilização e implantação embrionáriaDocumento8 páginasDescrevendo fertilização e implantação embrionáriaRenan VillacaAinda não há avaliações
- Gastrulação HumanaDocumento11 páginasGastrulação HumanaMaíra CaldeiraAinda não há avaliações
- Primeira A Terceira Semana de DesenvolvimentoDocumento6 páginasPrimeira A Terceira Semana de DesenvolvimentoHellen KathleenAinda não há avaliações
- Classificação dos óvulos e desenvolvimento embrionárioDocumento6 páginasClassificação dos óvulos e desenvolvimento embrionárioFelipe SantosAinda não há avaliações
- Da Terceira A Oitava SemanaDocumento45 páginasDa Terceira A Oitava Semanayasmim abreuAinda não há avaliações
- GastrulaçãoDocumento1 páginaGastrulaçãoarquivosanthonybioAinda não há avaliações
- Fecundação e Gestação PDFDocumento65 páginasFecundação e Gestação PDFLourenço Marques100% (3)
- Apresentação EmbriologiaDocumento46 páginasApresentação EmbriologiaEdson dos SantosAinda não há avaliações
- Bioligia 2Documento20 páginasBioligia 2Gabriel silvaAinda não há avaliações
- Biologia Do Desenvolvimento de MamíferosDocumento11 páginasBiologia Do Desenvolvimento de MamíferosGabi SoaresAinda não há avaliações
- OpenAccess Leitão 9788580391893 06Documento16 páginasOpenAccess Leitão 9788580391893 06isaac miraAinda não há avaliações
- EmbriogeneseDocumento20 páginasEmbriogenesemendesdecatricio2003Ainda não há avaliações
- Estudo sobre os principais eventos da embriogênese e fase fetalDocumento6 páginasEstudo sobre os principais eventos da embriogênese e fase fetalMariele Tesche KüsterAinda não há avaliações
- Origem do celoma e metameria nos animaisDocumento37 páginasOrigem do celoma e metameria nos animaisAndres RoberAinda não há avaliações
- 3 Semana de Desenvolvimento HumanoDocumento10 páginas3 Semana de Desenvolvimento HumanoStiven MoianaAinda não há avaliações
- SLIDE GASTRULAÇÃO, NEURULAÇÃO e ORGANOGÊNESEDocumento40 páginasSLIDE GASTRULAÇÃO, NEURULAÇÃO e ORGANOGÊNESEBiancaSantosAinda não há avaliações
- Citologia - Células DeciduaisDocumento2 páginasCitologia - Células DeciduaisAline GrazieleAinda não há avaliações
- Da Fecundação Ao NascimentoDocumento9 páginasDa Fecundação Ao NascimentoFaBeAnAinda não há avaliações
- Resumo EmbriologiaDocumento6 páginasResumo EmbriologiaMaria Helena Ribeiro LuvisaAinda não há avaliações
- Exercícios Desenvolvimento EmbrionárioDocumento2 páginasExercícios Desenvolvimento EmbrionárioIsione Ferreira100% (1)
- Linhagens de Organismos Metazoários 1 Autor João Miguel de Matos NogueiraDocumento14 páginasLinhagens de Organismos Metazoários 1 Autor João Miguel de Matos NogueiraJoão Victor FischerAinda não há avaliações
- Anatomia floral e formação do gametófitoDocumento13 páginasAnatomia floral e formação do gametófitoIsabella CapillaAinda não há avaliações
- Estudo dirigido parte 2: desenvolvimento embrionário e embriologiaDocumento7 páginasEstudo dirigido parte 2: desenvolvimento embrionário e embriologiacafé CharmosoAinda não há avaliações
- Atividade Divina 1Documento4 páginasAtividade Divina 1DannyMendesAinda não há avaliações
- Noções Embriologia emDocumento34 páginasNoções Embriologia emVon der WissenschaftAinda não há avaliações
- Trabalho Helmintologia VeterinariaDocumento8 páginasTrabalho Helmintologia VeterinariaPaulo JoseAinda não há avaliações
- Importância da água e nutrientes na dieta animalDocumento16 páginasImportância da água e nutrientes na dieta animalPaulo JoseAinda não há avaliações
- Influência de Diferentes Níveis de FDN Dietético No Consumo e Digestibilidade Aparente de Ovelhas Santa InêDocumento7 páginasInfluência de Diferentes Níveis de FDN Dietético No Consumo e Digestibilidade Aparente de Ovelhas Santa InêPaulo JoseAinda não há avaliações
- Mapa Mental Sistema Radicular de PlantasDocumento1 páginaMapa Mental Sistema Radicular de PlantasPaulo JoseAinda não há avaliações
- Mapa Mental Degradabilidade Ruminal Da Matéria SecaDocumento1 páginaMapa Mental Degradabilidade Ruminal Da Matéria SecaPaulo JoseAinda não há avaliações
- Avaliação do valor nutritivo de forrageiras de inverno em diferentes épocas de semeaduraDocumento1 páginaAvaliação do valor nutritivo de forrageiras de inverno em diferentes épocas de semeaduraPaulo JoseAinda não há avaliações
- Sistema digestório: como funciona a digestãoDocumento29 páginasSistema digestório: como funciona a digestãoOranceli Moreno Deganuti CarballoAinda não há avaliações
- Uma Nova Fórmula para o Cálculo de RaçãoDocumento8 páginasUma Nova Fórmula para o Cálculo de RaçãoPaulo JoseAinda não há avaliações
- Corpos Estranhos Esofágicos em CãesDocumento46 páginasCorpos Estranhos Esofágicos em CãesPaulo JoseAinda não há avaliações
- Anatomia abdominalDocumento22 páginasAnatomia abdominalBruno FuerstAinda não há avaliações
- Sistema Digestivo HumanoDocumento7 páginasSistema Digestivo HumanoPatricia CatalãoAinda não há avaliações
- 3 - ArgumentosDocumento16 páginas3 - ArgumentosCarina GasparAinda não há avaliações
- Enfermagem - UNIPLAN AULA Anatomia 06-10Documento20 páginasEnfermagem - UNIPLAN AULA Anatomia 06-10RosyAinda não há avaliações
- Teste 02Documento7 páginasTeste 02Célia Borges100% (1)
- Cap.17 Cordados Protocordados e VertebradosDocumento40 páginasCap.17 Cordados Protocordados e VertebradosChalu TamuraAinda não há avaliações
- Sistema Respirat Rio IDocumento22 páginasSistema Respirat Rio INeks Júnio Castro de CarvalhoAinda não há avaliações
- Tabela Comparativa Filos Do Reino AnimaliaDocumento1 páginaTabela Comparativa Filos Do Reino Animaliasergio.bsb100% (5)
- FI - Sistema Reprodutor Masculino e FemininoDocumento2 páginasFI - Sistema Reprodutor Masculino e FemininoTIHAinda não há avaliações
- Diferença Entre Sistema Reprodutor Masculino e FemininoDocumento8 páginasDiferença Entre Sistema Reprodutor Masculino e FemininoEurogénio RaúlAinda não há avaliações
- Apostila de Pronto SocorrismoDocumento103 páginasApostila de Pronto SocorrismoEmyster GaiaAinda não há avaliações
- Curso de yoga - Questionário de anatomia e fisiologiaDocumento3 páginasCurso de yoga - Questionário de anatomia e fisiologiaVicente MoliternoAinda não há avaliações
- Vter9 Fi Img 03Documento1 páginaVter9 Fi Img 03zavaAinda não há avaliações
- Aula Revisão NeuroanatomiaDocumento37 páginasAula Revisão NeuroanatomiaRoberta CostaAinda não há avaliações
- Noções gerais sobre células, imunidade, tecidos e sistemas osteo-articular e muscularDocumento27 páginasNoções gerais sobre células, imunidade, tecidos e sistemas osteo-articular e muscularPaula Fernandes67% (9)
- Teste de Conhecimento CitologiaDocumento6 páginasTeste de Conhecimento CitologiaMANIFESTO PORTO REALAinda não há avaliações
- Curso de Alquimia - Professor Olavo (1996)Documento83 páginasCurso de Alquimia - Professor Olavo (1996)Douglas LisboaAinda não há avaliações
- Os principais tipos de tecidos do corpo humanoDocumento27 páginasOs principais tipos de tecidos do corpo humanoSara CestariAinda não há avaliações
- Fichadetrabalho Sistema DigestivoDocumento3 páginasFichadetrabalho Sistema Digestivomaria100% (1)
- 6 Roteiro Sistema DigestórioDocumento2 páginas6 Roteiro Sistema Digestórioemelymorel2023Ainda não há avaliações
- Sistema MuscularDocumento10 páginasSistema MuscularjoaoAinda não há avaliações
- Ciclose celular em ElodeaDocumento3 páginasCiclose celular em ElodeaTássia SilveiraAinda não há avaliações
- Ossos Do Corpo HumanoDocumento4 páginasOssos Do Corpo HumanoDeyvid RabelloAinda não há avaliações
- Epitélio - Funções e ClassificaçãoDocumento3 páginasEpitélio - Funções e ClassificaçãoIzadora VeigaAinda não há avaliações
- 4 BaçoDocumento18 páginas4 BaçoMarcelly SeixasAinda não há avaliações
- Filo ArthropodaDocumento57 páginasFilo ArthropodaJosue Jesus SousaAinda não há avaliações
- Funções da peleDocumento2 páginasFunções da peleMárcia LeviAinda não há avaliações
- Sistema DigestivoDocumento3 páginasSistema DigestivoMarisa GonçalvesAinda não há avaliações
- Planos e Eixos AnatômicosDocumento11 páginasPlanos e Eixos AnatômicosTata PetroffAinda não há avaliações
- Sistema Circulatório LinfáticoDocumento17 páginasSistema Circulatório LinfáticoIsaac FloresAinda não há avaliações