Você também pode gostar
- No Limite de Seu Prazer - A DECI - Kira BaptistaDocumento319 páginasNo Limite de Seu Prazer - A DECI - Kira BaptistaLiliane Tumelero100% (4)
- Capt 5. Contribuições Das Neurociências À PsicopatologiaDocumento10 páginasCapt 5. Contribuições Das Neurociências À PsicopatologiaSara Fernandes ferreiraAinda não há avaliações
- Mapeamento e Diagnóstico Ambiental Da Trilha Da Pedra Do Machado Na Apa Da Serra Da Ibiapaba (Ce)Documento13 páginasMapeamento e Diagnóstico Ambiental Da Trilha Da Pedra Do Machado Na Apa Da Serra Da Ibiapaba (Ce)Flavio ZenAinda não há avaliações
- Mata Atlantica Fichas Das Areas PrioritariasDocumento879 páginasMata Atlantica Fichas Das Areas PrioritariasRavok100% (1)
- Questões Gordon AllportDocumento2 páginasQuestões Gordon AllportbiancaAinda não há avaliações
- Os Dominio Da Natureza Ab'saber Resenha 2003Documento7 páginasOs Dominio Da Natureza Ab'saber Resenha 2003Walena Marçal MagalhãesAinda não há avaliações
- ROSSI PANACHO ARASAKI 2009 Monitoramento Da Mastofauna Com ArmadilhasDocumento4 páginasROSSI PANACHO ARASAKI 2009 Monitoramento Da Mastofauna Com Armadilhasmarcelo.arasakiAinda não há avaliações
- Louro Cristina Et Al Investigação e Monitoria Das ACs Pomene. 2016Documento43 páginasLouro Cristina Et Al Investigação e Monitoria Das ACs Pomene. 2016Palmira AlfredoAinda não há avaliações
- PrevedelloCarvalho2006Pan BiogeografiaecerradoportuguesDocumento20 páginasPrevedelloCarvalho2006Pan BiogeografiaecerradoportuguesGabriel HenriqueAinda não há avaliações
- (Dissertação) Distribuição Atual de Onça-Parda (Puma Concolor) e Onça-Pintada (Panthera Onca) No Pantanal Brasileiro PDFDocumento29 páginas(Dissertação) Distribuição Atual de Onça-Parda (Puma Concolor) e Onça-Pintada (Panthera Onca) No Pantanal Brasileiro PDFReysane AlencarAinda não há avaliações
- O Lavrado Da Serra Da Lua em Roraima e Perspectivas para Estudos Da Herpetofauna Na RegiãoDocumento14 páginasO Lavrado Da Serra Da Lua em Roraima e Perspectivas para Estudos Da Herpetofauna Na Regiãopublischer100% (2)
- Perfis Ecologicos de Especies e Ecossistemas Costeiros em Mocambique DUNAS COSTEIRASDocumento32 páginasPerfis Ecologicos de Especies e Ecossistemas Costeiros em Mocambique DUNAS COSTEIRASEloi Carlos GoveAinda não há avaliações
- TCC ZoologiaDocumento22 páginasTCC ZoologiaReinaldo GomesAinda não há avaliações
- Plano de Manejo Arie Do BuritiDocumento34 páginasPlano de Manejo Arie Do BuritiMariana LampeAinda não há avaliações
- Atividades 7 Ano - 23Documento12 páginasAtividades 7 Ano - 23Karlene Pinheiro100% (1)
- 10 1016@j Jaridenv 2012 10 007Documento7 páginas10 1016@j Jaridenv 2012 10 007Fredson PereiraAinda não há avaliações
- Seleção de Áreas de Interesse Ecológico Através deDocumento8 páginasSeleção de Áreas de Interesse Ecológico Através deSurf AnalyticsAinda não há avaliações
- SILVEIRA Et Al. Prospecção Arqueológica em Áreas de Floresta, p.155-178 (Rev. MAE 09)Documento24 páginasSILVEIRA Et Al. Prospecção Arqueológica em Áreas de Floresta, p.155-178 (Rev. MAE 09)Elis OliveiraAinda não há avaliações
- Belarmina ProntoDocumento13 páginasBelarmina ProntoCllaudia FrançesAinda não há avaliações
- Brown Doodle Company Profile PresentationDocumento10 páginasBrown Doodle Company Profile PresentationThalia silvaAinda não há avaliações
- Protocolo AbilioDocumento22 páginasProtocolo AbilioAbilio Rafael MucaveleAinda não há avaliações
- ResenhaAbSber LimaSombraDocumento7 páginasResenhaAbSber LimaSombraJoao Paulo Araujo FerreiraAinda não há avaliações
- COUTINHO, R Costoes RochososDocumento102 páginasCOUTINHO, R Costoes RochososKarina100% (1)
- Ecologia de Onça-Pintada (Panthera Onca) Na CaatingaDocumento2 páginasEcologia de Onça-Pintada (Panthera Onca) Na Caatingamarcio vieiraAinda não há avaliações
- Abilio RafaelDocumento22 páginasAbilio RafaelAbilio Rafael MucaveleAinda não há avaliações
- Campanha de FaunaaDocumento19 páginasCampanha de FaunaaHiago Adamosky MachadoAinda não há avaliações
- AA-2013-0324. Macrofauna em CapoeirasDocumento14 páginasAA-2013-0324. Macrofauna em CapoeirasisabellashangresAinda não há avaliações
- Galoa Proceedings Epcc 79459Documento5 páginasGaloa Proceedings Epcc 79459hoteldonaanarececaoAinda não há avaliações
- Gray 2016Documento8 páginasGray 2016Guilherme D. Lauro DiasAinda não há avaliações
- Seminário Artigo EcotropDocumento16 páginasSeminário Artigo Ecotropdavid calazansAinda não há avaliações
- BryophytaDocumento8 páginasBryophytaWilliam PinheiroAinda não há avaliações
- 2017 SBG CTIG Paper 29Documento8 páginas2017 SBG CTIG Paper 29Agueda Jaime MilioneAinda não há avaliações
- DESAFIOS Dos Catadores de Caranguejos Na Reserva Extrativista Marinha Maracanã, Pará, Brasil.Documento12 páginasDESAFIOS Dos Catadores de Caranguejos Na Reserva Extrativista Marinha Maracanã, Pará, Brasil.Elizabeth SantosAinda não há avaliações
- BiogeografiaDocumento30 páginasBiogeografiaSebastião SantosAinda não há avaliações
- 74530-Texto Do Artigo-307080-1-10-20210817Documento17 páginas74530-Texto Do Artigo-307080-1-10-20210817Marcelo CampelloAinda não há avaliações
- Cortezzi Et Al.,2012 - Influência Da Ação Antrópica Sobre Macroinvertebrados de AssisDocumento8 páginasCortezzi Et Al.,2012 - Influência Da Ação Antrópica Sobre Macroinvertebrados de AssisRicardo Cardoso LeiteAinda não há avaliações
- Ilhas Vegetacionais Do Pantanal Matogrossense PDFDocumento3 páginasIlhas Vegetacionais Do Pantanal Matogrossense PDFJuniorShangresMedimAinda não há avaliações
- Caracterizaçào Empírica Da Fragilidade Ambiental Utilizando GeoprocessamentoDocumento9 páginasCaracterizaçào Empírica Da Fragilidade Ambiental Utilizando GeoprocessamentoAlmeida MucunaAinda não há avaliações
- Trabalho Final Pré ProjetoDocumento10 páginasTrabalho Final Pré Projetoevelinecunha23Ainda não há avaliações
- 3804-Texto Do Trabalho-6498-1-10-20190715Documento21 páginas3804-Texto Do Trabalho-6498-1-10-20190715Assocenio MaziveAinda não há avaliações
- BIOMAS CERRADO e PANTANALDocumento9 páginasBIOMAS CERRADO e PANTANALJéssica LimaAinda não há avaliações
- Areas Protegidas de MocambiqueDocumento16 páginasAreas Protegidas de MocambiqueMiguel Malua100% (2)
- As Florestas Secas Do Setor Meridional Da Serra Da Ibiapaba (Pi/ce) : Serviços Ecossistêmicos e ConservaçãoDocumento12 páginasAs Florestas Secas Do Setor Meridional Da Serra Da Ibiapaba (Pi/ce) : Serviços Ecossistêmicos e ConservaçãobicicleteirosemterraAinda não há avaliações
- Geodiversidade - Meio Abiotico PDFDocumento15 páginasGeodiversidade - Meio Abiotico PDFDercio100% (1)
- Fitogeografia: Mara Rojane Barros de MatosDocumento31 páginasFitogeografia: Mara Rojane Barros de MatosAlex WagnerAinda não há avaliações
- Artigo Projeto Salobo Versao EditorDocumento24 páginasArtigo Projeto Salobo Versao EditorSabrina SousaAinda não há avaliações
- Método de Marcação e Captura de Anfíbios - DiegoDocumento12 páginasMétodo de Marcação e Captura de Anfíbios - DiegoFátima SilvaAinda não há avaliações
- Ecologia Da AmazoniaDocumento71 páginasEcologia Da AmazoniaCarolina AmorimAinda não há avaliações
- Adequação Espacial e Conceitual para o Zoneamento Da APA Do PratigiDocumento10 páginasAdequação Espacial e Conceitual para o Zoneamento Da APA Do PratigiDanusa CamposAinda não há avaliações
- 03 Christiano Menezesetal AGIRASV2 N12010 FinalDocumento16 páginas03 Christiano Menezesetal AGIRASV2 N12010 FinalAnonymous YOfL2BE4Ainda não há avaliações
- ARTIGO Itororo FinalDocumento11 páginasARTIGO Itororo Finaldanusacampos5863Ainda não há avaliações
- 109 Sub 2080427802 Guia Repteis AnfibiosDocumento72 páginas109 Sub 2080427802 Guia Repteis AnfibioselsribeiroAinda não há avaliações
- Traduzido - Modeling Impacts of Climate Change On The Potential Habitat of An Endangered Brazilian Endemic Coral - Discussion About Deep Sea RefugiaDocumento24 páginasTraduzido - Modeling Impacts of Climate Change On The Potential Habitat of An Endangered Brazilian Endemic Coral - Discussion About Deep Sea RefugiarafislazzaAinda não há avaliações
- Hoehnea 36 2 T 2Documento16 páginasHoehnea 36 2 T 2Canal Alessa Pereira RosaAinda não há avaliações
- 1997forestedgegrassland InsectesSociaux MajeretalDocumento13 páginas1997forestedgegrassland InsectesSociaux Majeretalvt.lima.2018Ainda não há avaliações
- Ictiofauna Da Reserva Nacional de ChimanDocumento56 páginasIctiofauna Da Reserva Nacional de ChimanAgueda Jaime MilioneAinda não há avaliações
- Modelo de Diagnostico AmbientalDocumento39 páginasModelo de Diagnostico Ambientalleruaites100% (1)
- Capacidade de Carga e Análise Da Efetividade Da Revitalização de Uma Trilha Interpretativa Na RPPN Uru - Lapa/PRDocumento12 páginasCapacidade de Carga e Análise Da Efetividade Da Revitalização de Uma Trilha Interpretativa Na RPPN Uru - Lapa/PRRegino Raul ComarAinda não há avaliações
- Marquesetal Parintins CBCS0181Documento6 páginasMarquesetal Parintins CBCS0181brissa senraAinda não há avaliações
- Twenty Years of Land Cover Change in The Southeastern Peruvian Amazon - Implications For Biodiversity Conservation PDFDocumento14 páginasTwenty Years of Land Cover Change in The Southeastern Peruvian Amazon - Implications For Biodiversity Conservation PDFRyan MarquesAinda não há avaliações
- Dinâmica e Estoque de Carbono em Florestas Primárias Na Região de Manaus-AMDocumento6 páginasDinâmica e Estoque de Carbono em Florestas Primárias Na Região de Manaus-AMcamilamilhomem5Ainda não há avaliações
- Parque Natural Municipal Rio Do Peixe, Joaçaba, Santa CatarinaNo EverandParque Natural Municipal Rio Do Peixe, Joaçaba, Santa CatarinaAinda não há avaliações
- Eucalyptus Camaldulensis É Uma: Árvore AustráliaDocumento2 páginasEucalyptus Camaldulensis É Uma: Árvore AustráliaSavio SitoeAinda não há avaliações
- Faculdade de Ciencias Direcção Académica Comprovativo de InscriçãoDocumento1 páginaFaculdade de Ciencias Direcção Académica Comprovativo de InscriçãoSavio SitoeAinda não há avaliações
- Relatório 3 BDDocumento4 páginasRelatório 3 BDSavio SitoeAinda não há avaliações
- Relatório 3 BDDocumento4 páginasRelatório 3 BDSavio SitoeAinda não há avaliações
- Aula 1 Pratica BD 2023Documento8 páginasAula 1 Pratica BD 2023Savio SitoeAinda não há avaliações
- OvogéneseDocumento2 páginasOvogéneseSavio SitoeAinda não há avaliações
- Simulado 1 - FuvestDocumento25 páginasSimulado 1 - FuvestGiovanna Galvão ScheideggerAinda não há avaliações
- Resultados Do Programa Monitorização Lixo Marinho em Praias 2020 Rev 1 - 0Documento22 páginasResultados Do Programa Monitorização Lixo Marinho em Praias 2020 Rev 1 - 0Daniel CarapetoAinda não há avaliações
- Artigo Desenvolvimento Na Primeira Infância - Perspectiva de René SpitzDocumento8 páginasArtigo Desenvolvimento Na Primeira Infância - Perspectiva de René SpitzAntonio Augusto Pinto JuniorAinda não há avaliações
- Plasticidade Cerebral 1 1Documento20 páginasPlasticidade Cerebral 1 1thamiresdhellenAinda não há avaliações
- Ensinando Criancas A Amar - FinalDocumento198 páginasEnsinando Criancas A Amar - FinalCabelo dos Sonhos100% (1)
- Sos - Simulado Enem - 2º DiaDocumento45 páginasSos - Simulado Enem - 2º Diacardosoyasmim838Ainda não há avaliações
- Ensaio FilosóficoDocumento6 páginasEnsaio FilosóficoPutz R4sput1nAinda não há avaliações
- PLANO DA ZELHANIR E ALINE (1) (Zelhanir Fatima Rodrigues Furtado)Documento11 páginasPLANO DA ZELHANIR E ALINE (1) (Zelhanir Fatima Rodrigues Furtado)Whagna Rodrigues DuarteAinda não há avaliações
- Calendário de Postagens 365 DiasDocumento13 páginasCalendário de Postagens 365 Diaspolyana welida majevskiAinda não há avaliações
- Reino Fungi - EXERCICIOS - RespondidosDocumento6 páginasReino Fungi - EXERCICIOS - Respondidosbeth rodiguesAinda não há avaliações
- Relatório de Levantamento MaterialDocumento2 páginasRelatório de Levantamento MaterialHeltonsubrg SystemAinda não há avaliações
- Palestra o Poder Da Autorresponsabilidade v20 06 22Documento46 páginasPalestra o Poder Da Autorresponsabilidade v20 06 22Cosme Gomes100% (2)
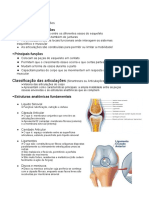
- ArtrologiaDocumento4 páginasArtrologiaJosé GuimarãesAinda não há avaliações
- Uma Vida em AzulDocumento15 páginasUma Vida em AzulVicente TavaresAinda não há avaliações
- 01 Estudo Dirigido Do Filo PoriferaDocumento5 páginas01 Estudo Dirigido Do Filo PoriferaBernardo JacobsenAinda não há avaliações
- Genetica ENEMDocumento3 páginasGenetica ENEMDudeckAinda não há avaliações
- 1.cinética Microbiana 13Documento43 páginas1.cinética Microbiana 13davi100% (1)
- Livro Neuropsicopedagogia Do TOD. Transtorno Opositor Desafiador-1Documento75 páginasLivro Neuropsicopedagogia Do TOD. Transtorno Opositor Desafiador-1llatyffaAinda não há avaliações
- Avaliação - GENETICADocumento5 páginasAvaliação - GENETICAAgrícola MK - Fenix cereaisAinda não há avaliações
- MAPA EM 2ano V2 Ciencias Da Natureza PFDocumento55 páginasMAPA EM 2ano V2 Ciencias Da Natureza PFMarcãoquimico MarquesAinda não há avaliações
- Exigências Climáticas para A Produção de Milho No PiauíDocumento4 páginasExigências Climáticas para A Produção de Milho No PiauíGil BatistaAinda não há avaliações
- Desmistificando - MKUltraDocumento82 páginasDesmistificando - MKUltraGabriel SantosAinda não há avaliações
- ReZero - LN 29Documento372 páginasReZero - LN 29vitor vlogs e jogos brasilAinda não há avaliações
- Fator ImpactoDocumento15 páginasFator ImpactojorgethiagoAinda não há avaliações
- Respiracao Do Sol Hashira HandbookDocumento6 páginasRespiracao Do Sol Hashira Handbookrebekacc12Ainda não há avaliações
- AvencaDocumento4 páginasAvencaRac A BruxaAinda não há avaliações
- Desenvolvimento II Cronograma 23.1 Sexta FeiraDocumento9 páginasDesenvolvimento II Cronograma 23.1 Sexta FeiraJeniffer PereiraAinda não há avaliações