Você também pode gostar
- Autobiografia de um polvo: e outras narrativas de antecipaçãoNo EverandAutobiografia de um polvo: e outras narrativas de antecipaçãoAinda não há avaliações
- Ornithorhynchus. Juntamente Com As: Mamífero Austrália. Ornithorhynchidae Equidnas Monotremados Ovíparos MonotípicaDocumento7 páginasOrnithorhynchus. Juntamente Com As: Mamífero Austrália. Ornithorhynchidae Equidnas Monotremados Ovíparos MonotípicaChronoSyAinda não há avaliações
- Mosquitos assassinos: arbovírus e arbovirosesNo EverandMosquitos assassinos: arbovírus e arbovirosesAinda não há avaliações
- Ap3 DeuterosDocumento2 páginasAp3 DeuterosYan Romano MonteiroAinda não há avaliações
- Lista de Zoologia ResolvidaDocumento5 páginasLista de Zoologia ResolvidaluisAinda não há avaliações
- Aves Do Período Cretáceo Da Era MesozoicaDocumento4 páginasAves Do Período Cretáceo Da Era MesozoicaMario Arthur FavrettoAinda não há avaliações
- As últimas notícias do Sapiens: Uma revolução nas nossas origensNo EverandAs últimas notícias do Sapiens: Uma revolução nas nossas origensNota: 5 de 5 estrelas5/5 (1)
- Nome:: Soma: 30Documento4 páginasNome:: Soma: 30Francisco WesleyAinda não há avaliações
- Vertebrados Amniotas - SauropsidaDocumento76 páginasVertebrados Amniotas - SauropsidagloriambenAinda não há avaliações
- 10 Eeq 60 16Documento5 páginas10 Eeq 60 16lagartojrAinda não há avaliações
- Diversidade Biologica Dos Deuterostomados Aula 20 Volume3Documento19 páginasDiversidade Biologica Dos Deuterostomados Aula 20 Volume3Pedro Henrique Marques dos SantosAinda não há avaliações
- Invertebrados PDFDocumento13 páginasInvertebrados PDFDiego VazAinda não há avaliações
- A Origem Das AvesDocumento16 páginasA Origem Das AvesAntonio ManucciAinda não há avaliações
- 4 Sarcopterygii Conquista Do Meio Terrestre AmphibiaDocumento28 páginas4 Sarcopterygii Conquista Do Meio Terrestre Amphibiaabilio eliasAinda não há avaliações
- Cronologia Da Evolução HumanaDocumento12 páginasCronologia Da Evolução HumanaTAG100% (1)
- Biologia 3Documento11 páginasBiologia 3fernandalinsAinda não há avaliações
- Aula 1 Introdução A PaleontologiaDocumento5 páginasAula 1 Introdução A PaleontologiaJoaquina AndressaAinda não há avaliações
- A Origem Das AvesDocumento5 páginasA Origem Das AvesnathaliaAinda não há avaliações
- Origem e Evolucao Do Plano Corpóreo de Gastropodes 01Documento11 páginasOrigem e Evolucao Do Plano Corpóreo de Gastropodes 01Vanessa Bezerra100% (3)
- Portfólio em Grupo - Análise Evolutiva de Dois Grupos de VertebradosDocumento8 páginasPortfólio em Grupo - Análise Evolutiva de Dois Grupos de VertebradosRê RodriguesAinda não há avaliações
- Revolução Dos Bichos-1Documento18 páginasRevolução Dos Bichos-1Anna LessaAinda não há avaliações
- 5 Os Aminiota RépteisDocumento33 páginas5 Os Aminiota Répteisabilio eliasAinda não há avaliações
- Aula 3 Entomologia ForenseDocumento5 páginasAula 3 Entomologia Forenseluana diasAinda não há avaliações
- Origem e Evolução Dos VertebradosDocumento98 páginasOrigem e Evolução Dos Vertebradosjokercorvo0% (3)
- Hexapoda - TagmasDocumento50 páginasHexapoda - TagmasAndre LuisAinda não há avaliações
- Evidências Da EvoluçãoDocumento5 páginasEvidências Da EvoluçãoValdinei IsacAinda não há avaliações
- Aula 1Documento11 páginasAula 1mateus silvaAinda não há avaliações
- Questões Artrópodes Médio/DifícilDocumento98 páginasQuestões Artrópodes Médio/DifícilAna Beatriz DiasAinda não há avaliações
- Origem e Irradiação de Tetrapoda PDFDocumento24 páginasOrigem e Irradiação de Tetrapoda PDFWilliam RafaelAinda não há avaliações
- 13 Evolução de Primeiros Vertebrados Voadores 4 Ano VictoriaDocumento10 páginas13 Evolução de Primeiros Vertebrados Voadores 4 Ano VictoriaSadique Sadique AtiboAinda não há avaliações
- Evolução Dos Vertebrados 05-10-2019 Pré-VestibularDocumento17 páginasEvolução Dos Vertebrados 05-10-2019 Pré-Vestibularidelfonso filhoAinda não há avaliações
- Paleontologia - Apostila - Fósseis e TafonomiaDocumento26 páginasPaleontologia - Apostila - Fósseis e TafonomiaClaudia Costa100% (1)
- Artrópode TraduzidoDocumento13 páginasArtrópode TraduzidoMarlla AroucheAinda não há avaliações
- 09 OrthopteraDocumento71 páginas09 OrthopteraMorena VàsconezAinda não há avaliações
- Exercício - CópiaDocumento6 páginasExercício - CópiaBrenno VarjãoAinda não há avaliações
- Exercícios Sobre Evolução 2022Documento6 páginasExercícios Sobre Evolução 2022Lussandra MarquezAinda não há avaliações
- Provas AnterioresDocumento16 páginasProvas AnterioresBruno Mian SilvaAinda não há avaliações
- Estudo Dirigido de AmniotasDocumento2 páginasEstudo Dirigido de AmniotasTatto AraujoAinda não há avaliações
- Soria S. 1975.ciclos de Vida Dos Polinizadores Do Cacaueiro Forcipomyia SPP PDFDocumento20 páginasSoria S. 1975.ciclos de Vida Dos Polinizadores Do Cacaueiro Forcipomyia SPP PDFA.JAinda não há avaliações
- Evolucionismo e EspeciaçãoDocumento12 páginasEvolucionismo e Especiaçãomagary_mgAinda não há avaliações
- I UNIDADE - Possíveis Questões Zoologia VDocumento6 páginasI UNIDADE - Possíveis Questões Zoologia VAna Paula Gomes BispoAinda não há avaliações
- Introdução A PaleontologiaDocumento73 páginasIntrodução A PaleontologiaJessyka Abreu Chagas100% (4)
- 2004 (UFPR - Terceiro Dia) Biologia, Física e Química.Documento11 páginas2004 (UFPR - Terceiro Dia) Biologia, Física e Química.jubileu100% (1)
- Questões Comentadas de EvoluçãoDocumento6 páginasQuestões Comentadas de EvoluçãoFelipe Almeida PinhoAinda não há avaliações
- Macroevolução Dos AnimaisDocumento3 páginasMacroevolução Dos AnimaisInês MauadAinda não há avaliações
- 700 Questões de BiologiaDocumento573 páginas700 Questões de BiologiaGiovani Milla Chonps BiologiaAinda não há avaliações
- Agnathas Atuais e ChondrichthyesDocumento30 páginasAgnathas Atuais e ChondrichthyesMarcia LobAinda não há avaliações
- 6 Aves PDFDocumento30 páginas6 Aves PDFBruno DiorgenesAinda não há avaliações
- Pterosauros No BrasilDocumento6 páginasPterosauros No BrasilIracy MaianyAinda não há avaliações
- A Origem Das Aves: Uma Discussão ContínuaDocumento2 páginasA Origem Das Aves: Uma Discussão ContínuaMario Arthur FavrettoAinda não há avaliações
- AlgasDocumento39 páginasAlgasVictor Sousa100% (1)
- Atividade CORDADOSDocumento3 páginasAtividade CORDADOSJuliano Abreu PrattiAinda não há avaliações
- MArDINA (Guardado Automaticamente) - 1Documento19 páginasMArDINA (Guardado Automaticamente) - 1Rafael faruqueAinda não há avaliações
- T. Rex Quando As Galinhas Tinham Dentes Autor Pavilhão Do ConhecimentoDocumento19 páginasT. Rex Quando As Galinhas Tinham Dentes Autor Pavilhão Do ConhecimentoGabriel MarinoAinda não há avaliações
- EC - 7º Ano - Ciências - Animais VertebradosDocumento11 páginasEC - 7º Ano - Ciências - Animais VertebradosRosana ValeAinda não há avaliações
- Vocacional 2 Formiga Teste de Avaliação Modulo 8Documento4 páginasVocacional 2 Formiga Teste de Avaliação Modulo 8Isabel FortunatoAinda não há avaliações
- Aula 21Documento15 páginasAula 21Pedro Henrique Marques dos SantosAinda não há avaliações
- Evidências Morfo-Ecológicas Acerca Da Origem Das Aves A Partir Dos TheropodaDocumento4 páginasEvidências Morfo-Ecológicas Acerca Da Origem Das Aves A Partir Dos TheropodaLetícia CavalcantiAinda não há avaliações
- Trab de Geogra de Urban TERMINADODocumento15 páginasTrab de Geogra de Urban TERMINADOAcacioMonjaneAinda não há avaliações
- Saeb9ano - 2simuladoDocumento2 páginasSaeb9ano - 2simuladoAdriane LopesAinda não há avaliações
- Admin, COMPORTAMENTO DE CONSUMO ECOLOGICAMENTE CONSCIENTEDocumento28 páginasAdmin, COMPORTAMENTO DE CONSUMO ECOLOGICAMENTE CONSCIENTEMarques Da DeolindaAinda não há avaliações
- Universidade Federal de Santa Catarina Programa de Pós-Graduação em Engenharia AmbientalDocumento133 páginasUniversidade Federal de Santa Catarina Programa de Pós-Graduação em Engenharia AmbientalLOGAN LOGANAinda não há avaliações
- SEMAS - Instrução Normativa No 50, de 25 de Agosto de 2010Documento4 páginasSEMAS - Instrução Normativa No 50, de 25 de Agosto de 2010Jefferson Bittencourt VenâncioAinda não há avaliações
- Republica Brasileira em 1890 .: Ensaio Do BrasilDocumento345 páginasRepublica Brasileira em 1890 .: Ensaio Do BrasilAlex RegisAinda não há avaliações
- (Regulamentação) DECRETO #15.798 - Mato Grosso Do Sul (2021)Documento16 páginas(Regulamentação) DECRETO #15.798 - Mato Grosso Do Sul (2021)Ariane MoraesAinda não há avaliações
- 6 TERRAPLENAGEM - ATERROS - NeDocumento11 páginas6 TERRAPLENAGEM - ATERROS - NeRicardo LimaAinda não há avaliações
- Aleitedebarros, Clicia Julie Batista BarataDocumento21 páginasAleitedebarros, Clicia Julie Batista BarataDelano MaiaAinda não há avaliações
- Monografia ARMANDO AZEVEDODocumento48 páginasMonografia ARMANDO AZEVEDOAermando Azevedo CastigoAinda não há avaliações
- Ofício Condicionante - Cer. Serido - Rev 03 RJDocumento3 páginasOfício Condicionante - Cer. Serido - Rev 03 RJRivaldo Nóbrega JuniorAinda não há avaliações
- Unidade 1 OutorgaDocumento39 páginasUnidade 1 OutorgaJaque RibeiroAinda não há avaliações
- Ciências Do AmbienteDocumento97 páginasCiências Do AmbienteBruna SilveiraAinda não há avaliações
- Zampollo - Neder-Iniciativas - de Produção - Mais - Limpa - Na - Indústria - de - Óleo - e - GásDocumento10 páginasZampollo - Neder-Iniciativas - de Produção - Mais - Limpa - Na - Indústria - de - Óleo - e - GásTardan TardanAinda não há avaliações
- Lei Nº 20.922, de 16 de Outubro de 2013Documento45 páginasLei Nº 20.922, de 16 de Outubro de 2013Fábio GomesAinda não há avaliações
- Coraclean Ed: FISPQ-Ficha de Informação de Segurança de Produtos Químicos em Acordo Com A NBR-14725Documento5 páginasCoraclean Ed: FISPQ-Ficha de Informação de Segurança de Produtos Químicos em Acordo Com A NBR-14725Fernando GiacominiAinda não há avaliações
- AO2 Prova Meio Ambiente Sociedade e CidadaniaDocumento14 páginasAO2 Prova Meio Ambiente Sociedade e Cidadaniaizabelliburgarelli11Ainda não há avaliações
- 3 - ARTIGO - Diálogo Social e Agenda 2030 - Maria Hemília Fonseca - 2020Documento22 páginas3 - ARTIGO - Diálogo Social e Agenda 2030 - Maria Hemília Fonseca - 2020Gustavo De Carvalho ArgoloAinda não há avaliações
- Portfólio Efotec SolarDocumento12 páginasPortfólio Efotec SolarSimone LimaAinda não há avaliações
- Fatores BioticosDocumento17 páginasFatores BioticosSEAinda não há avaliações
- Luc 220 - FispqDocumento10 páginasLuc 220 - Fispqjulioebc2014Ainda não há avaliações
- 10760Documento3 páginas10760Rádio Cariri HitzAinda não há avaliações
- Aula 3 - Efe - Matriz EnergéticaDocumento15 páginasAula 3 - Efe - Matriz EnergéticaJúniorLopesAinda não há avaliações
- Exame - Q4Documento4 páginasExame - Q4GetlowAinda não há avaliações
- Dimensionamento de Reservatório de Amortecimento - ExemploDocumento31 páginasDimensionamento de Reservatório de Amortecimento - ExemploRodrigoAinda não há avaliações
- 8º Parte 1 - Fontes e Tipos de EnergiaDocumento6 páginas8º Parte 1 - Fontes e Tipos de EnergiaMaíra MonteiroAinda não há avaliações
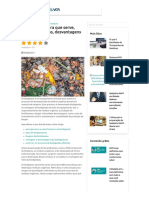
- Biodigestor - para Que Serve, Tipos, Vantagens, Desvantagens - VG ResíduosDocumento8 páginasBiodigestor - para Que Serve, Tipos, Vantagens, Desvantagens - VG ResíduosDoze DozautoAinda não há avaliações
- Piscicultura No Estado Do Par - Situao Atual e Perspectivas - 2014Documento7 páginasPiscicultura No Estado Do Par - Situao Atual e Perspectivas - 2014pedro bulhoesAinda não há avaliações
- Atividades 5º Ano - 8º QuinzenaDocumento11 páginasAtividades 5º Ano - 8º QuinzenaMarco LawrenzAinda não há avaliações
- Port. Basico - ZambelliDocumento72 páginasPort. Basico - ZambellibrunoAinda não há avaliações
- Análise linguística: Afinal a que se refere?No EverandAnálise linguística: Afinal a que se refere?Ainda não há avaliações
- A Trama do conhecimento: Teoria, método e escrita em ciência e pesquisaNo EverandA Trama do conhecimento: Teoria, método e escrita em ciência e pesquisaAinda não há avaliações
- Ecologia de riachos de montanha da Mata AtlânticaNo EverandEcologia de riachos de montanha da Mata AtlânticaNota: 5 de 5 estrelas5/5 (1)
- Como passar na OAB 1ª Fase: direito constitucional: 400 questões comentadasNo EverandComo passar na OAB 1ª Fase: direito constitucional: 400 questões comentadasNota: 5 de 5 estrelas5/5 (1)
- Cartas xamânicas: A descoberta do poder através da energia dos animaisNo EverandCartas xamânicas: A descoberta do poder através da energia dos animaisNota: 4 de 5 estrelas4/5 (10)
- Fitoenergética: A Energia das Plantas no Equilíbrio da AlmaNo EverandFitoenergética: A Energia das Plantas no Equilíbrio da AlmaNota: 5 de 5 estrelas5/5 (17)
- Dieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNo EverandDieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNota: 5 de 5 estrelas5/5 (4)
- Ativações Espirituais: Obsessão e Evolução pelos Implantes ExtrafísicosNo EverandAtivações Espirituais: Obsessão e Evolução pelos Implantes ExtrafísicosNota: 4.5 de 5 estrelas4.5/5 (18)
- Práticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNo EverandPráticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNota: 4 de 5 estrelas4/5 (1)
- Biossíntese, Funções e Aplicações dos Metabólitos Secundários de PlantasNo EverandBiossíntese, Funções e Aplicações dos Metabólitos Secundários de PlantasAinda não há avaliações
- Como passar na OAB 1ª Fase: direito penal: 325 questões comentadasNo EverandComo passar na OAB 1ª Fase: direito penal: 325 questões comentadasAinda não há avaliações
- Como passar na OAB 1ª Fase: direito do trabalho: 230 questões comentadasNo EverandComo passar na OAB 1ª Fase: direito do trabalho: 230 questões comentadasNota: 5 de 5 estrelas5/5 (1)
- Serviços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesNo EverandServiços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesAinda não há avaliações
- Criança e a Experiência Afetiva com a NaturezaNo EverandCriança e a Experiência Afetiva com a NaturezaAinda não há avaliações
- Universo quântico e sincronicidade. A visão antrópica. As coincidências significativas. O inconsciente coletivo. O papel das pandemias no caminho evolutivo humano.No EverandUniverso quântico e sincronicidade. A visão antrópica. As coincidências significativas. O inconsciente coletivo. O papel das pandemias no caminho evolutivo humano.Nota: 5 de 5 estrelas5/5 (1)
- Questões de linguística aplicada ao ensino: da teoria à práticaNo EverandQuestões de linguística aplicada ao ensino: da teoria à práticaNota: 5 de 5 estrelas5/5 (1)
- Série Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNo EverandSérie Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNota: 5 de 5 estrelas5/5 (1)
- Estranhas coincidências em sua vida. Pequenos eventos curiosos. Pressentimentos. Telepatia. Isso acontece com você também? A física quântica e a teoria da sincronicidade explicam os fenômenos extra-seNo EverandEstranhas coincidências em sua vida. Pequenos eventos curiosos. Pressentimentos. Telepatia. Isso acontece com você também? A física quântica e a teoria da sincronicidade explicam os fenômenos extra-seNota: 4 de 5 estrelas4/5 (2)
- O homem, único "bicho" contraversor das leis da naturezaNo EverandO homem, único "bicho" contraversor das leis da naturezaAinda não há avaliações
- A Teoria do Domínio do Fato e o Autor Imediato nos Crimes Contra a Administração PúblicaNo EverandA Teoria do Domínio do Fato e o Autor Imediato nos Crimes Contra a Administração PúblicaAinda não há avaliações
- A Psicanálise da História: A teoria relevada dos quatro elementosNo EverandA Psicanálise da História: A teoria relevada dos quatro elementosAinda não há avaliações
- Águas de Rio Branco: Dos Lençóis Freáticos às TorneirasNo EverandÁguas de Rio Branco: Dos Lençóis Freáticos às TorneirasAinda não há avaliações