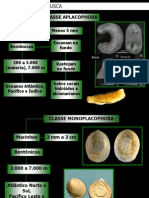
Você também pode gostar
- TEXTO Origem CordadosDocumento6 páginasTEXTO Origem CordadosdeyffersonAinda não há avaliações
- Introdução Aos EcdysozoaDocumento2 páginasIntrodução Aos EcdysozoaLourdes Elmoor LoureiroAinda não há avaliações
- Introdução Ao LophotrochozoaDocumento14 páginasIntrodução Ao LophotrochozoaDione Queiroz Amorim100% (1)
- Anatomia Na TaxonomiaDocumento5 páginasAnatomia Na TaxonomiaDanilo José Lima De SousaAinda não há avaliações
- 12044403042012botanica Sistematica e Economica Aula 6Documento27 páginas12044403042012botanica Sistematica e Economica Aula 6JOAO VICTOR LAUBER BATISTAAinda não há avaliações
- Relações Filogeneticas BilateriaDocumento5 páginasRelações Filogeneticas BilateriaMayara LimaAinda não há avaliações
- Origem-Evolução e Filogenia de Chordata e CraniataDocumento27 páginasOrigem-Evolução e Filogenia de Chordata e CraniataMarcia LobAinda não há avaliações
- Resumo CordadosDocumento5 páginasResumo CordadosMarcia MelloAinda não há avaliações
- Estudo Dirigido 1Documento2 páginasEstudo Dirigido 1Ericca LoboAinda não há avaliações
- Aula 5Documento6 páginasAula 5mateus silvaAinda não há avaliações
- Ornithorhynchus. Juntamente Com As: Mamífero Austrália. Ornithorhynchidae Equidnas Monotremados Ovíparos MonotípicaDocumento7 páginasOrnithorhynchus. Juntamente Com As: Mamífero Austrália. Ornithorhynchidae Equidnas Monotremados Ovíparos MonotípicaChronoSyAinda não há avaliações
- Aula 21Documento15 páginasAula 21Pedro Henrique Marques dos SantosAinda não há avaliações
- Go Tarefa Semanal 3 Serie 5e7a66a9146bbDocumento4 páginasGo Tarefa Semanal 3 Serie 5e7a66a9146bbAnderson Adolfo ScipiãoAinda não há avaliações
- USP 06 - Arthropoda 1Documento30 páginasUSP 06 - Arthropoda 1Célio Moura Neto100% (2)
- DivEvoPlan - Caracterização Das AngiospermasDocumento23 páginasDivEvoPlan - Caracterização Das AngiospermasNelson AlencarAinda não há avaliações
- Evidências Da EvoluçãoDocumento5 páginasEvidências Da EvoluçãoValdinei IsacAinda não há avaliações
- Estudo Dirigido de AmniotasDocumento2 páginasEstudo Dirigido de AmniotasTatto AraujoAinda não há avaliações
- Sobre A Nova Proposta para As Relações Filogenéticas Dos Principais Grupos de Dinossauros e As Implicações em Sua Origem e DiversificaçãoDocumento8 páginasSobre A Nova Proposta para As Relações Filogenéticas Dos Principais Grupos de Dinossauros e As Implicações em Sua Origem e DiversificaçãoJoão EduardoAinda não há avaliações
- Origem e Irradiação de Tetrapoda PDFDocumento24 páginasOrigem e Irradiação de Tetrapoda PDFWilliam RafaelAinda não há avaliações
- 6 Aves PDFDocumento30 páginas6 Aves PDFBruno DiorgenesAinda não há avaliações
- Exercicios Artropodes, Moluscos e ArtropodesDocumento2 páginasExercicios Artropodes, Moluscos e ArtropodesReydner PaulaAinda não há avaliações
- Cronologia Da Evolução HumanaDocumento12 páginasCronologia Da Evolução HumanaTAG100% (1)
- Invertebrados PDFDocumento13 páginasInvertebrados PDFDiego VazAinda não há avaliações
- 3.09.ET.O Registro Fossilífero e o Processo Evolutivo - Megabiotas ExtintasDocumento25 páginas3.09.ET.O Registro Fossilífero e o Processo Evolutivo - Megabiotas ExtintasProfessor BiologiaAinda não há avaliações
- Teoria Pseudo PDFDocumento10 páginasTeoria Pseudo PDFDiana MendoncaAinda não há avaliações
- Hexapoda - TagmasDocumento50 páginasHexapoda - TagmasAndre LuisAinda não há avaliações
- Agnathas Atuais e ChondrichthyesDocumento30 páginasAgnathas Atuais e ChondrichthyesMarcia LobAinda não há avaliações
- Argumentos Do EvolucionismoDocumento9 páginasArgumentos Do EvolucionismoHélder WafungaAinda não há avaliações
- Trabalho de Biologia Evolutiva FichaDocumento4 páginasTrabalho de Biologia Evolutiva FichaGo Cuambe OrlandoAinda não há avaliações
- Origem Das Asas Dos InsetosDocumento9 páginasOrigem Das Asas Dos InsetosMyllena BastosAinda não há avaliações
- 4 VetebradosDocumento62 páginas4 VetebradosAmanda OliveiraAinda não há avaliações
- 4 Vetebrados PDFDocumento62 páginas4 Vetebrados PDFLuiza FormigaAinda não há avaliações
- FICHA de Leitura BioDocumento5 páginasFICHA de Leitura BioJeremias ManhiqueAinda não há avaliações
- Visão Geral Das Descobertas e Classificação Dos Terópodes Não AvianosDocumento92 páginasVisão Geral Das Descobertas e Classificação Dos Terópodes Não AvianosJoão EduardoAinda não há avaliações
- Análise Cladística de Oxalis Sect. Thamnoxys ARTDocumento12 páginasAnálise Cladística de Oxalis Sect. Thamnoxys ARTcarlosAinda não há avaliações
- Filogenia 2Documento9 páginasFilogenia 2PabloDarlanAinda não há avaliações
- Aprofundamento-Biologia-Construindo Cladogramas-27-09-2023Documento11 páginasAprofundamento-Biologia-Construindo Cladogramas-27-09-2023Mauricio SantanaAinda não há avaliações
- Exercício ProtozoologiaDocumento11 páginasExercício ProtozoologiaDenner SouzaAinda não há avaliações
- Resumo Zoo. ChordataDocumento9 páginasResumo Zoo. ChordataLalaAinda não há avaliações
- I UNIDADE - Possíveis Questões Zoologia VDocumento6 páginasI UNIDADE - Possíveis Questões Zoologia VAna Paula Gomes BispoAinda não há avaliações
- Código Internacional de Nomeclatura Zoológica PDFDocumento27 páginasCódigo Internacional de Nomeclatura Zoológica PDFElained SilvaAinda não há avaliações
- Aula 4 Euangiospermas:: Introdução Às MonocotiledôneasDocumento9 páginasAula 4 Euangiospermas:: Introdução Às MonocotiledôneasJean SantosAinda não há avaliações
- Filo ArthropodaDocumento5 páginasFilo ArthropodaIslane SantosAinda não há avaliações
- Angiospermas ClassificaçãoDocumento32 páginasAngiospermas ClassificaçãoRhuan CarvalhoAinda não há avaliações
- Estudo Dirigido de Biologia Os Segundos Anos 1 BimestreDocumento3 páginasEstudo Dirigido de Biologia Os Segundos Anos 1 BimestreArthur FiaminiAinda não há avaliações
- Evolucao Gonçalves Orlando Cuambe CuambeDocumento7 páginasEvolucao Gonçalves Orlando Cuambe CuambeGo Cuambe OrlandoAinda não há avaliações
- Apostila ArthropodaDocumento35 páginasApostila Arthropodawendellupton8035Ainda não há avaliações
- Origem e Evolução Dos VertebradosDocumento98 páginasOrigem e Evolução Dos Vertebradosjokercorvo0% (3)
- 09 - Taxonomia X Cladística - Sistemática Filogenética.Documento20 páginas09 - Taxonomia X Cladística - Sistemática Filogenética.Marcelo Loer Bellini Monjardim BarbozaAinda não há avaliações
- Classificação Dos Seres VivosDocumento68 páginasClassificação Dos Seres VivosMaria IzabelAinda não há avaliações
- Aula de Nomenclatura e Coleções ZoológicasDocumento77 páginasAula de Nomenclatura e Coleções ZoológicasEduardo Got'z100% (2)
- Resenha Sobre Os "Filos" Pogonophora, Echiura e Sipuncula e As Relações Filogenéticas Com AnnelidaDocumento2 páginasResenha Sobre Os "Filos" Pogonophora, Echiura e Sipuncula e As Relações Filogenéticas Com AnnelidaPietra BarbosaAinda não há avaliações
- Os AgnathaDocumento57 páginasOs AgnathaKAÍS RODRIGUES DE NOVAESAinda não há avaliações
- MonocotiledôneasDocumento34 páginasMonocotiledôneasDaniela IvanAinda não há avaliações
- Plano de Aula Zoo2 ArthropodaDocumento7 páginasPlano de Aula Zoo2 ArthropodaEdimara DelerinoAinda não há avaliações
- Neves, Rocha - Evolução HumanaDocumento28 páginasNeves, Rocha - Evolução HumanadadacestmoiAinda não há avaliações
- 09 OrthopteraDocumento71 páginas09 OrthopteraMorena VàsconezAinda não há avaliações
- Autobiografia de um polvo: e outras narrativas de antecipaçãoNo EverandAutobiografia de um polvo: e outras narrativas de antecipaçãoAinda não há avaliações
- Resumo Dos Filos - Platelmintos, Nematelmintos, Equinodermos, Artrópode, Molusco, AnelídeoDocumento6 páginasResumo Dos Filos - Platelmintos, Nematelmintos, Equinodermos, Artrópode, Molusco, Anelídeopacola10100% (4)
- Noturno Arthropoda ArtropodizaçãoDocumento34 páginasNoturno Arthropoda ArtropodizaçãoDavid Da Silva ConstantinoAinda não há avaliações
- PlatelmintosDocumento23 páginasPlatelmintosMel LunaAinda não há avaliações
- Aula - PLATYHELMINTES e NEMATODA PDFDocumento58 páginasAula - PLATYHELMINTES e NEMATODA PDFDeboraAinda não há avaliações
- Ciemcias 7º Ano 2Documento3 páginasCiemcias 7º Ano 2IreneAguiarAinda não há avaliações
- Especialidade - MoluscosDocumento18 páginasEspecialidade - MoluscosSanderson HenriqueAinda não há avaliações
- Filo MoluscosDocumento19 páginasFilo MoluscosDenilson JosemirAinda não há avaliações
- Ecologia e Concha MolluscaDocumento42 páginasEcologia e Concha MolluscaManoel NetoAinda não há avaliações
- Resumo Reino Animal 1Documento2 páginasResumo Reino Animal 1Michel Stórquio BelmiroAinda não há avaliações
- Relatório de Aula Prática AnelídeosDocumento8 páginasRelatório de Aula Prática AnelídeosMarcos AntônioAinda não há avaliações
- Centopéias Ou Lacraias e Piolho de CobraDocumento2 páginasCentopéias Ou Lacraias e Piolho de CobraFilhoAinda não há avaliações
- Mollusca CefalopodosDocumento61 páginasMollusca CefalopodosLeonardo HalfenAinda não há avaliações
- 03 Estudo Dirigido Do Filo PlatyhelminthesDocumento4 páginas03 Estudo Dirigido Do Filo PlatyhelminthesBernardo JacobsenAinda não há avaliações
- Platelmintos e NematódeosDocumento15 páginasPlatelmintos e NematódeosIrídia BarbosaAinda não há avaliações
- TABELA Fisiologia Animal Comparada Cassiano BIO 03Documento1 páginaTABELA Fisiologia Animal Comparada Cassiano BIO 03Renan DisnerAinda não há avaliações
- Branch I UraDocumento6 páginasBranch I UraBriza RochaAinda não há avaliações
- Aula 2-4 Chaetognatha CycloneuraliaDocumento92 páginasAula 2-4 Chaetognatha CycloneuraliaHanna KarolinaAinda não há avaliações
- Filo MolluscaDocumento14 páginasFilo MolluscaMiranda AdrianoAinda não há avaliações
- Exercício de Ciências 7º AnoDocumento3 páginasExercício de Ciências 7º AnoBárbara PereiraAinda não há avaliações
- Texto LofoforadosDocumento3 páginasTexto LofoforadosFelipe Micali NuvoloniAinda não há avaliações
- Aula EcdysozoaDocumento39 páginasAula EcdysozoaCaro RecaldeAinda não há avaliações
- Bio2 Roteiro 03Documento3 páginasBio2 Roteiro 03biowellAinda não há avaliações
- Introdução Ao Estudo Do Filo MolluscaDocumento17 páginasIntrodução Ao Estudo Do Filo Molluscajon.alves100% (1)
- AnelideosDocumento9 páginasAnelideosJair Ferreira LimaAinda não há avaliações
- Prova Cie 4bim 7anoDocumento2 páginasProva Cie 4bim 7anoNivaldo JúniorAinda não há avaliações
- Invertebrados: Tabela ComparativaDocumento1 páginaInvertebrados: Tabela ComparativaRebeca Maciel MoreiraAinda não há avaliações
- Pan Arthropod A Resumo (Modo de CompatibilidadeDocumento49 páginasPan Arthropod A Resumo (Modo de CompatibilidadeTiago CostaAinda não há avaliações
- Bio 2 EstdirigidoDocumento2 páginasBio 2 EstdirigidogiselestevesAinda não há avaliações
- Filo AnnelidaDocumento16 páginasFilo AnnelidaLuana100% (1)
- Atividade 2º AnoDocumento2 páginasAtividade 2º AnoAna Paula SilveiraAinda não há avaliações