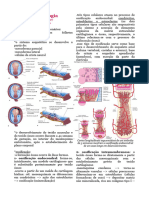
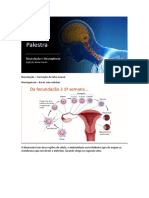
Você também pode gostar
- Gastrulação em HumanosDocumento11 páginasGastrulação em HumanosMaíra CaldeiraAinda não há avaliações
- Período Embrionário Ou de Organogênese EstendeDocumento5 páginasPeríodo Embrionário Ou de Organogênese EstendeRafaela Marques FerreiraAinda não há avaliações
- Embriologia - Esqueleto Axial (Resumo)Documento10 páginasEmbriologia - Esqueleto Axial (Resumo)hina kurigayaAinda não há avaliações
- Estudo Dirigido Parte 2Documento7 páginasEstudo Dirigido Parte 2café CharmosoAinda não há avaliações
- Terceira Semana de DesenvolvimentoDocumento11 páginasTerceira Semana de Desenvolvimentomararaujo2664Ainda não há avaliações
- ED - Fisio Embrio p2Documento4 páginasED - Fisio Embrio p2Felipe RomãoAinda não há avaliações
- IFB - Embriologia IIDocumento10 páginasIFB - Embriologia IIEmanuel NatanaelAinda não há avaliações
- Zoologia GeralDocumento13 páginasZoologia GeralGo Cuambe OrlandoAinda não há avaliações
- GastrulaçãoDocumento2 páginasGastrulaçãoJulia XimenesAinda não há avaliações
- SomitosDocumento3 páginasSomitosRafael TeixeiraAinda não há avaliações
- ED Biomorfo PR2 GabaritoDocumento5 páginasED Biomorfo PR2 Gabaritomaria vitoriaAinda não há avaliações
- Arquiv 1504Documento6 páginasArquiv 1504Kaline CristinaAinda não há avaliações
- Morfoooo ApreseDocumento2 páginasMorfoooo ApreseAna CauanaAinda não há avaliações
- GastrulaçãoDocumento1 páginaGastrulaçãoarquivosanthonybioAinda não há avaliações
- Terceira Semana Do Desenvolvimnto EmbrionárioDocumento7 páginasTerceira Semana Do Desenvolvimnto EmbrionárioJefferssonh AssysAinda não há avaliações
- Embriologia - 3 Semana PDFDocumento26 páginasEmbriologia - 3 Semana PDFFrança OliveiraAinda não há avaliações
- Roteiro 1 EmbriologiaDocumento8 páginasRoteiro 1 EmbriologiaraulAinda não há avaliações
- Esqueleto Axial-Grupo 1Documento17 páginasEsqueleto Axial-Grupo 1Mario RivasAinda não há avaliações
- Resumão Do Corujão - Embrio - Prova 2 - 1) Gastrulação (Aula 1)Documento6 páginasResumão Do Corujão - Embrio - Prova 2 - 1) Gastrulação (Aula 1)jairofioraAinda não há avaliações
- EMBRIOLOGIADocumento9 páginasEMBRIOLOGIAViarmano D Claudina FirminoAinda não há avaliações
- Aula 3 - Embriologia Do Sistema Nervoso AuraDocumento22 páginasAula 3 - Embriologia Do Sistema Nervoso AuraFERNANDO GONCALVES PEREIRAAinda não há avaliações
- Embriogênese Veterinária - Desenvolvimento Sistema Digestivo BovinoDocumento3 páginasEmbriogênese Veterinária - Desenvolvimento Sistema Digestivo BovinoPaulo JoseAinda não há avaliações
- Aula 3. Gastrulacao - 3 SemanaDocumento43 páginasAula 3. Gastrulacao - 3 SemanaAuryelle Sousa AzevedoAinda não há avaliações
- Gastrulao e Organogenese 2Documento38 páginasGastrulao e Organogenese 2Geise Vasconcelos100% (1)
- Unidade 1 - Neurociencia Do ComportamentoDocumento13 páginasUnidade 1 - Neurociencia Do ComportamentoAndré SantanaAinda não há avaliações
- Divisao Da MitoseDocumento11 páginasDivisao Da MitoseAnonymous rgPNr8Xa7Ainda não há avaliações
- Módulo de Concepção-Tutoria 5-Bebê de FasesDocumento22 páginasMódulo de Concepção-Tutoria 5-Bebê de FasesMatheus UrsiAinda não há avaliações
- Ciclo Básico: Embriologia Do Sistema MuscularDocumento4 páginasCiclo Básico: Embriologia Do Sistema MuscularFidelis JuniorAinda não há avaliações
- ResumoDocumento5 páginasResumoRafaela Marques FerreiraAinda não há avaliações
- UntitledDocumento25 páginasUntitledBruno Souza FonsecaAinda não há avaliações
- Resumo Da Terceira Semana Do Desenvolvimento EmbrionárioDocumento2 páginasResumo Da Terceira Semana Do Desenvolvimento EmbrionárioClaudia Nunes100% (1)
- 3 SEMANA (GASTRULACAO) .EditDocumento11 páginas3 SEMANA (GASTRULACAO) .EditrosangelaAinda não há avaliações
- Continuacao Da Gastrulacao (Somitos, Celoma)Documento8 páginasContinuacao Da Gastrulacao (Somitos, Celoma)rosangelaAinda não há avaliações
- Resumo de HistologiaDocumento8 páginasResumo de HistologiaClaudia_ymAinda não há avaliações
- NEURULAÇÃODocumento9 páginasNEURULAÇÃOKauane TedescoAinda não há avaliações
- EMBRIOLOGIA Trabalho para EntregarDocumento7 páginasEMBRIOLOGIA Trabalho para Entregarandressa dos SantosAinda não há avaliações
- 3 Semana de Desenvolvimento HumanoDocumento10 páginas3 Semana de Desenvolvimento HumanoStiven MoianaAinda não há avaliações
- SandraDocumento9 páginasSandraAntonio AnastacioAinda não há avaliações
- Tutoria 2 - Embriologia Do SNDocumento3 páginasTutoria 2 - Embriologia Do SNAna Paula Viégas100% (1)
- Em Briolo GiDocumento12 páginasEm Briolo GivincentAinda não há avaliações
- 3 J 4 - 8 Semanas Do Desenvolvimento EmbrionárioDocumento10 páginas3 J 4 - 8 Semanas Do Desenvolvimento EmbrionárioBixete Med Assis LauraAinda não há avaliações
- Aula 4. NeurulacaoDocumento26 páginasAula 4. NeurulacaoAuryelle Sousa AzevedoAinda não há avaliações
- Desenvolvimento Do EncéfaloDocumento6 páginasDesenvolvimento Do EncéfaloGabriel MartinsAinda não há avaliações
- Principais Eventos Da Segunda e Terceira Semana Do Desenvolvimento EmbrionárioDocumento6 páginasPrincipais Eventos Da Segunda e Terceira Semana Do Desenvolvimento EmbrionárioPaolaRaeski100% (1)
- Estudo Dirigido de EmbriologiaDocumento6 páginasEstudo Dirigido de EmbriologiaMariele Tesche KüsterAinda não há avaliações
- ESTUDO DIRIGIDO 6 - EmbriologiaDocumento4 páginasESTUDO DIRIGIDO 6 - EmbriologiaCarla Caroline Alves De LimaAinda não há avaliações
- Desenvolvimento Cranio Facial - OdontologiaDocumento7 páginasDesenvolvimento Cranio Facial - OdontologiaBruna Goncalves De OliveiraAinda não há avaliações
- Terceira SemanaDocumento2 páginasTerceira SemanaMoniely GonçalvesAinda não há avaliações
- Capítulo 1-Embriologia Do Sistema MusculoesqueléticoDocumento4 páginasCapítulo 1-Embriologia Do Sistema MusculoesqueléticoLuiz CarlosAinda não há avaliações
- Embriologia - Seminario Desenvolv. ProtocordadosDocumento36 páginasEmbriologia - Seminario Desenvolv. ProtocordadosJoão Luiz Pereira JúniorAinda não há avaliações
- Palestra - Embriologia Do Sistema NervosoDocumento17 páginasPalestra - Embriologia Do Sistema NervosoKas PrintAinda não há avaliações
- 62bc6110dfc0d 95 Embriologia Embriologia Dos MembrosDocumento9 páginas62bc6110dfc0d 95 Embriologia Embriologia Dos Membrosmaysa landimAinda não há avaliações
- 3 Semana de DesenvolvimentoDocumento5 páginas3 Semana de DesenvolvimentoVicttoria Melo100% (1)
- Diferenciação Dos Folhetos Embrionarios - SanarDocumento9 páginasDiferenciação Dos Folhetos Embrionarios - SanarMel na MeddAinda não há avaliações
- Atividade Embriologia Sistemas Cartilaginoso e ÓsseoDocumento3 páginasAtividade Embriologia Sistemas Cartilaginoso e Ósseoluana andradeAinda não há avaliações
- EmbriologiaDocumento10 páginasEmbriologialouren maurinAinda não há avaliações
- Embriologia IIDocumento3 páginasEmbriologia IIMarina BarbieriAinda não há avaliações
- Segunda Semana Do Desenvolvimento EmbrionárioDocumento5 páginasSegunda Semana Do Desenvolvimento EmbrionárioJefferssonh AssysAinda não há avaliações
- Transtornos Psiquiátricos Relacionados A Gestação e Ao Puerpério. - Compressed - Compressed - CompressedDocumento42 páginasTranstornos Psiquiátricos Relacionados A Gestação e Ao Puerpério. - Compressed - Compressed - CompressedRaissa QuintanilhaAinda não há avaliações
- Especificidades Anatômicas Do RN - EMERGENCIALDocumento89 páginasEspecificidades Anatômicas Do RN - EMERGENCIALRaissa QuintanilhaAinda não há avaliações
- Uploads - Manual Do Professor 2020Documento32 páginasUploads - Manual Do Professor 2020Raissa QuintanilhaAinda não há avaliações
- Disforia Pós PartoDocumento7 páginasDisforia Pós PartoRaissa QuintanilhaAinda não há avaliações
- Declaração de Artigo No PreloDocumento1 páginaDeclaração de Artigo No PreloRoberta CantarelaAinda não há avaliações
- Gaiolas e Asas - Rubem Alves 01 PDFDocumento2 páginasGaiolas e Asas - Rubem Alves 01 PDFJoãoSunahAinda não há avaliações
- Doe 5813 23032021Documento33 páginasDoe 5813 23032021John BlessedAinda não há avaliações
- Corpo e Ancestralidade Fabio LimaDocumento14 páginasCorpo e Ancestralidade Fabio LimaDenise David CaxiasAinda não há avaliações
- Curso 222309 Aula 05 5d72 CompletoDocumento33 páginasCurso 222309 Aula 05 5d72 CompletoCamila da SilveiraAinda não há avaliações
- Memoria Descritiva - Cobertura Metalica - COLEPDocumento60 páginasMemoria Descritiva - Cobertura Metalica - COLEPstar_starAinda não há avaliações
- Zine - Organizando Um Coletivo Feminista - Um Guia Pratico - GarrafeministaDocumento19 páginasZine - Organizando Um Coletivo Feminista - Um Guia Pratico - GarrafeministaCarolina SoaresAinda não há avaliações
- Modelo Ação RescisóriaDocumento5 páginasModelo Ação RescisóriaBianca Caroline da SilvaAinda não há avaliações
- Cópia de Receituário Especial para ImpressãoDocumento1 páginaCópia de Receituário Especial para ImpressãoLetícia MeloAinda não há avaliações
- Avaliação de Ciências - 5º Ano - 2022 - 4º BimestreDocumento6 páginasAvaliação de Ciências - 5º Ano - 2022 - 4º Bimestrecmazzoni_3100% (10)
- 4a Aula Síncrona Leis de NewtonDocumento64 páginas4a Aula Síncrona Leis de NewtonJorge twttAinda não há avaliações
- Paper 2Documento7 páginasPaper 2Aline SalcidesAinda não há avaliações
- Curso Técnico em Refrigeração e Climatização 1200h - Cetec AraguaínaDocumento79 páginasCurso Técnico em Refrigeração e Climatização 1200h - Cetec AraguaínaJorge ChavesAinda não há avaliações
- A Influência Dos Youtubers Na Vida Dos AdolescentesDocumento1 páginaA Influência Dos Youtubers Na Vida Dos AdolescentesThuany GomesAinda não há avaliações
- Passo A Passo IT Registro e Atendimento FalhaDocumento13 páginasPasso A Passo IT Registro e Atendimento Falhajefferson gomesAinda não há avaliações
- Tabela EL30 Techaccus1Documento3 páginasTabela EL30 Techaccus1Lisiane da silvaAinda não há avaliações
- A Crise de Integridade - Warren W. WiersbeDocumento58 páginasA Crise de Integridade - Warren W. WiersbeAllan Acioly100% (1)
- Adicionando Hachuras Personalizadas No AutoCAD PDFDocumento2 páginasAdicionando Hachuras Personalizadas No AutoCAD PDFValter AraujoAinda não há avaliações
- Revolução Cubana - Prof. AlbanirDocumento14 páginasRevolução Cubana - Prof. AlbanirAlbanir FaleirosAinda não há avaliações
- Captura de Tela 2021-07-27 À(s) 14.36.53Documento64 páginasCaptura de Tela 2021-07-27 À(s) 14.36.53tamara carvalhoAinda não há avaliações
- UA 04 - Introdução Às Capacidades (Skills)Documento11 páginasUA 04 - Introdução Às Capacidades (Skills)Susana CastroAinda não há avaliações
- A Fina Lâmina Da Palavra PDFDocumento30 páginasA Fina Lâmina Da Palavra PDFJoão NetoAinda não há avaliações
- Cinco Ritos TibetanosDocumento5 páginasCinco Ritos Tibetanoscarros-120Ainda não há avaliações
- Autor Da Minha Vida - Sérgio Lopes - LETRAS - MusDocumento4 páginasAutor Da Minha Vida - Sérgio Lopes - LETRAS - MusmesaxmelAinda não há avaliações
- Kevin Reed - Membresia de IgrejaDocumento3 páginasKevin Reed - Membresia de IgrejaJessica Juntocom MiqueiasAinda não há avaliações
- Ficha BladesingerDocumento3 páginasFicha BladesingerLuis PalosoAinda não há avaliações
- O Congresso de Milão HQ LetrariaDocumento45 páginasO Congresso de Milão HQ LetrariaQueli Dornelles MoraisAinda não há avaliações
- Cartilha BandeiradeLutas 2019versaofinalDocumento18 páginasCartilha BandeiradeLutas 2019versaofinalbrenda cruzAinda não há avaliações
- Operador de Videomonitoramento PDFDocumento28 páginasOperador de Videomonitoramento PDFMarcelo Fonseca100% (2)
- A Arca de NoéDocumento10 páginasA Arca de NoéElisangela SantiagoAinda não há avaliações
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Distúrbios internos da articulação temporomandibular para clínicos e especialistas: Anatomia, patologia e diagnóstico diferencialNo EverandDistúrbios internos da articulação temporomandibular para clínicos e especialistas: Anatomia, patologia e diagnóstico diferencialNota: 5 de 5 estrelas5/5 (1)
- Comunicação de más notícias: a distância entre morte encefálica e a doação de órgãoNo EverandComunicação de más notícias: a distância entre morte encefálica e a doação de órgãoAinda não há avaliações
- Rejuvenescer Com O Plasma Sanguíneo Dos JovensNo EverandRejuvenescer Com O Plasma Sanguíneo Dos JovensAinda não há avaliações
- História da Anatomia Através da Dissecação do Corpo HumanoNo EverandHistória da Anatomia Através da Dissecação do Corpo HumanoAinda não há avaliações