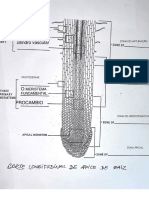
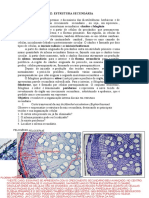
Você também pode gostar
- Apostila de ManicureDocumento32 páginasApostila de ManicureMari Dias75% (8)
- Desagregacao e DepuracaoDocumento40 páginasDesagregacao e Depuracaojosuepedroso_866688967% (3)
- Análise de Resistência Mecânica de Terminal de Eixo Cardan - Trabalho Referencia PDFDocumento90 páginasAnálise de Resistência Mecânica de Terminal de Eixo Cardan - Trabalho Referencia PDFGuilherme NascimentoAinda não há avaliações
- Projeto Balança Aerodinamica para Tunel de Vento FEAUDocumento8 páginasProjeto Balança Aerodinamica para Tunel de Vento FEAUCrystianne LilianAinda não há avaliações
- 7 - 8 Polaridade em Estacas de PlantasDocumento2 páginas7 - 8 Polaridade em Estacas de PlantasArnaldo ChivamboAinda não há avaliações
- Tecido VascularDocumento64 páginasTecido VascularEUNICE MACEDOAinda não há avaliações
- Fitormônios Prof GunaroDocumento1 páginaFitormônios Prof GunarogunarodeoliveiraAinda não há avaliações
- BiologiaDocumento4 páginasBiologiaMarta OliveiraAinda não há avaliações
- O Efeito Da Polaridade Na Formação de Raízes e Dos Rebentos Novos Num Ramo Cortado de Hibiscus.Documento7 páginasO Efeito Da Polaridade Na Formação de Raízes e Dos Rebentos Novos Num Ramo Cortado de Hibiscus.Alberto César MachavaAinda não há avaliações
- Unidade XV PDFDocumento39 páginasUnidade XV PDFBrunaCleitonAinda não há avaliações
- Resumo - FitohormôniosDocumento14 páginasResumo - FitohormôniosMarcelo PadovaniAinda não há avaliações
- Anatomia e Fisiologia de Peixes de Agua Doce Aplicada A PisciculturaDocumento46 páginasAnatomia e Fisiologia de Peixes de Agua Doce Aplicada A Pisciculturarob casAinda não há avaliações
- E - Aula 1 Transporte de FloemaDocumento39 páginasE - Aula 1 Transporte de FloemaDelivelton Alexandre LongarayAinda não há avaliações
- Hormônios ApostilaDocumento22 páginasHormônios ApostilaTroy SilvaAinda não há avaliações
- Aula II Cordados IDocumento44 páginasAula II Cordados ICarlos ToffoloAinda não há avaliações
- Botânica 2022 Enem - 102127Documento47 páginasBotânica 2022 Enem - 102127Evellyn MouraAinda não há avaliações
- Motilidade GastrointestinalDocumento70 páginasMotilidade GastrointestinalElla Vieira100% (1)
- FloemaDocumento31 páginasFloemaLaís BarretoAinda não há avaliações
- Fisio RespostasDocumento3 páginasFisio RespostasThaysa VieiraAinda não há avaliações
- Tabela HormoniosDocumento1 páginaTabela HormoniosNatalia Carvalho Fabricio da Silva100% (1)
- Bve 3009 AuxinasDocumento59 páginasBve 3009 AuxinasLuisa Fernanda RodríguezAinda não há avaliações
- Hormonios e Movimento VegetalDocumento10 páginasHormonios e Movimento Vegetalcamila warmelingAinda não há avaliações
- Cap 5Documento20 páginasCap 5Jocarlos AlbertonAinda não há avaliações
- Glossario PDFDocumento40 páginasGlossario PDFleo1221Ainda não há avaliações
- Biologia 4Documento11 páginasBiologia 4Joana StylesAinda não há avaliações
- Anatomia e Fisiologia de Peixes de Agua Doce Aplicada A PisciculturaDocumento44 páginasAnatomia e Fisiologia de Peixes de Agua Doce Aplicada A PisciculturaDalila CotaAinda não há avaliações
- Estudo Dirigido (AV2)Documento3 páginasEstudo Dirigido (AV2)Areshiya KurivueAinda não há avaliações
- ProtozoáriosDocumento26 páginasProtozoáriosThiago Almeida de Carvalho100% (1)
- Botanica Evolucao Das PlantasDocumento51 páginasBotanica Evolucao Das PlantasJean BrilhanteAinda não há avaliações
- 9 Aula - Sistema - Circulatorio - Dos - InsetosDocumento2 páginas9 Aula - Sistema - Circulatorio - Dos - InsetosRodrigo EstevesAinda não há avaliações
- Fisiologia VegetalDocumento7 páginasFisiologia VegetalAnne CarolineAinda não há avaliações
- 2a Prova BotDocumento5 páginas2a Prova BotGabrielle TeixeiraAinda não há avaliações
- Sabatina 3Documento14 páginasSabatina 3Marcos MotaAinda não há avaliações
- Grupo AschelminthesDocumento3 páginasGrupo AschelminthesAnne Judiz Casteiro de MeloAinda não há avaliações
- Resumo - Heteroconta DiatomaceasDocumento3 páginasResumo - Heteroconta DiatomaceasBreno Nunes da Silva OliveiraAinda não há avaliações
- Agua e MembranaDocumento36 páginasAgua e MembranaMarcio CastroAinda não há avaliações
- Um Pouco de DisciplinasDocumento173 páginasUm Pouco de DisciplinasJesus PedroAinda não há avaliações
- Reino ProtoctistaDocumento46 páginasReino ProtoctistagianAinda não há avaliações
- SanarFlix - Bomba de Sódio e Potássio (Mapa Mental)Documento1 páginaSanarFlix - Bomba de Sódio e Potássio (Mapa Mental)Gabriel Elias100% (1)
- Filo Composto Por Minhocas e Sanguessugas e Vermes MarinhosDocumento16 páginasFilo Composto Por Minhocas e Sanguessugas e Vermes MarinhosNatalia MafraAinda não há avaliações
- Filo AnnelidaDocumento4 páginasFilo AnnelidaAnne Judiz Casteiro de MeloAinda não há avaliações
- Reino VegetalDocumento53 páginasReino VegetalYasmin Vitoria BertonciniAinda não há avaliações
- Sistemas Circulatórios Dos Animais (Comparação) - Cola Da WebDocumento1 páginaSistemas Circulatórios Dos Animais (Comparação) - Cola Da WebAviva LimaAinda não há avaliações
- Aula - Transporte de Água e Fotoassimildaos - COMPLETADocumento34 páginasAula - Transporte de Água e Fotoassimildaos - COMPLETAotacide RosarioAinda não há avaliações
- Aula BryophytaDocumento33 páginasAula BryophytaKarla AraújoAinda não há avaliações
- Transporte No FloemaDocumento32 páginasTransporte No FloemaAirton Jose Trento FilhoAinda não há avaliações
- Zoologia AgrícolaDocumento46 páginasZoologia AgrícolaIndiana SantosAinda não há avaliações
- QueliceradoDocumento4 páginasQueliceradoDhenifer PedreirasAinda não há avaliações
- Cap 05Documento31 páginasCap 05lauramquintal27Ainda não há avaliações
- Resumo Artropodes + Prova CsaDocumento9 páginasResumo Artropodes + Prova CsaInês MauadAinda não há avaliações
- Fisiologia Das Angiospermas 1Documento20 páginasFisiologia Das Angiospermas 1Otavio AugustoAinda não há avaliações
- Resumo Fisiologia VegetaDocumento6 páginasResumo Fisiologia VegetaJane BuzolicAinda não há avaliações
- Relacoes UterofetaisDocumento2 páginasRelacoes Uterofetaisgabriella pachecoAinda não há avaliações
- Resumo BveDocumento3 páginasResumo BveBárbara CarolinaAinda não há avaliações
- 2 Resumo Transportes Membrana Celular 2020 21Documento13 páginas2 Resumo Transportes Membrana Celular 2020 21Laura MonteiroAinda não há avaliações
- Aula RaizDocumento21 páginasAula RaizIrapuã JuniorAinda não há avaliações
- Biofísica - Membrana PlasmáticaDocumento3 páginasBiofísica - Membrana PlasmáticaGabriela JobAinda não há avaliações
- Transdução de SinaisDocumento6 páginasTransdução de SinaisthuanesgAinda não há avaliações
- Reguladores de Crescimento FV AvancadaDocumento37 páginasReguladores de Crescimento FV AvancadaLuiz Felipe SiqueiraAinda não há avaliações
- IófitasDocumento6 páginasIófitasOsnei RibeiroAinda não há avaliações
- 6 Algas Protistas 2Documento85 páginas6 Algas Protistas 2Leonardo LealAinda não há avaliações
- Transportes Pela MPDocumento3 páginasTransportes Pela MPLeticia Buratto100% (1)
- Heterokotophyta PHAEOPHYCEAEDocumento4 páginasHeterokotophyta PHAEOPHYCEAEBreno Nunes da Silva OliveiraAinda não há avaliações
- CephalochordataDocumento18 páginasCephalochordataBEATRIZ GOMES RAMOSAinda não há avaliações
- Origem Dos Gnathostomata e ChondrichyesDocumento157 páginasOrigem Dos Gnathostomata e ChondrichyesBEATRIZ GOMES RAMOSAinda não há avaliações
- Caracteristicas Anatomicas e Morfologicas Da Estrutura Foliar PDFDocumento13 páginasCaracteristicas Anatomicas e Morfologicas Da Estrutura Foliar PDFmaites miguelAinda não há avaliações
- p5 CopiarDocumento6 páginasp5 CopiarBeatrizAinda não há avaliações
- Pratica4 CopiarprontoDocumento4 páginasPratica4 CopiarprontoBEATRIZ GOMES RAMOSAinda não há avaliações
- Pratica2.2-Copiar-Pronto P EntregaDocumento2 páginasPratica2.2-Copiar-Pronto P EntregaBEATRIZ GOMES RAMOSAinda não há avaliações
- Prática 4Documento8 páginasPrática 4BEATRIZ GOMES RAMOSAinda não há avaliações
- Metabissulfito de SódioDocumento1 páginaMetabissulfito de SódioEdileuson MedeirosAinda não há avaliações
- Sistema Agua GeladaDocumento22 páginasSistema Agua GeladaElias Barros de FigueiredoAinda não há avaliações
- 05 V2 - Colunas - Ficha - de - Apontamentos - 11a - ClasseDocumento25 páginas05 V2 - Colunas - Ficha - de - Apontamentos - 11a - ClasseJoao Dique Maguichire PkayAinda não há avaliações
- Introdução A Quimica AnalíticaDocumento5 páginasIntrodução A Quimica AnalíticaGuilherme FernandesAinda não há avaliações
- Escoamento de Fluidos IncompressíveisDocumento42 páginasEscoamento de Fluidos IncompressíveisEnegia CafemaniaAinda não há avaliações
- Medicação em Enfermagem Pediatrica - Jéssica - 2sem2022Documento44 páginasMedicação em Enfermagem Pediatrica - Jéssica - 2sem2022CatarinaAinda não há avaliações
- Bases NitrogenadasDocumento3 páginasBases NitrogenadasLais MarquesAinda não há avaliações
- Exercícios Resolvidos de Química PDFDocumento140 páginasExercícios Resolvidos de Química PDFgiovanna200467% (3)
- Gas ToxinDocumento6 páginasGas ToxinWalter GuimarãesAinda não há avaliações
- Relatório Da PilhaDocumento8 páginasRelatório Da PilhasoniaAinda não há avaliações
- Catalise BifuncionalDocumento26 páginasCatalise BifuncionalDaniel Pedrosa LopesAinda não há avaliações
- AstaxantinaDocumento2 páginasAstaxantinacaducampAinda não há avaliações
- Direito Quântico Goffredo Telles PDFDocumento25 páginasDireito Quântico Goffredo Telles PDFbbraga_rühleAinda não há avaliações
- Artigo Lauren Zauza - Eugenia UnifloraDocumento13 páginasArtigo Lauren Zauza - Eugenia UnifloraLauren ZauzaAinda não há avaliações
- Aula 1 - Introdução À FarmacologiaDocumento106 páginasAula 1 - Introdução À FarmacologiaTais Takeda100% (1)
- Informativo Tecnico #6 SiteDocumento13 páginasInformativo Tecnico #6 SiteJorge RosalAinda não há avaliações
- Trabalho de PqiDocumento12 páginasTrabalho de PqiMauro RomeroAinda não há avaliações
- Capitulo 4Documento27 páginasCapitulo 4Francisco MachadoAinda não há avaliações
- FT5 Mole 2022 CorreçãoDocumento3 páginasFT5 Mole 2022 Correçãojonas47Ainda não há avaliações
- Caderno Completo de EPIDocumento37 páginasCaderno Completo de EPIAnonymous ERRDz7uVAinda não há avaliações
- Procedimento Coleta Solo PDFDocumento8 páginasProcedimento Coleta Solo PDFMarcelo ZawadzkiAinda não há avaliações
- Metodologia de Fracionamento Da MOSDocumento7 páginasMetodologia de Fracionamento Da MOSLucasAinda não há avaliações
- 198 - Análise Bromatológica de Sorvete FormuladoDocumento4 páginas198 - Análise Bromatológica de Sorvete FormuladoAna Beatriz CorrêaAinda não há avaliações
- Resolução ANP N. 45-2014 (Especificção Do Biodiesel)Documento9 páginasResolução ANP N. 45-2014 (Especificção Do Biodiesel)Everaldo LimaAinda não há avaliações
- 1 AVALIAÇÃO DE CIÊNCIAS 9º Ano - 1º BIMESTRE 2023Documento5 páginas1 AVALIAÇÃO DE CIÊNCIAS 9º Ano - 1º BIMESTRE 2023Felipe TavaresAinda não há avaliações
- MODULO 3 BiologiaDocumento146 páginasMODULO 3 BiologiaMARINA NhambeAinda não há avaliações
- 2 Bed 6 CDocumento2 páginas2 Bed 6 CLucialdo d'ArêdeAinda não há avaliações