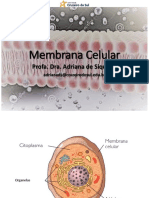
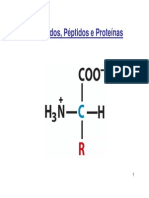
Você também pode gostar
- BIOLOGIA 13Documento8 páginasBIOLOGIA 13samstudyyyAinda não há avaliações
- Membrana plasmática e seus componentesDocumento13 páginasMembrana plasmática e seus componentesChristian FarinaAinda não há avaliações
- Membrana Plasmática - Características e Resumo - Mundo EducaçãoDocumento3 páginasMembrana Plasmática - Características e Resumo - Mundo EducaçãoMarcos Vinícius Neves OlsanAinda não há avaliações
- Estudo APG 02 - Celulas ..Documento10 páginasEstudo APG 02 - Celulas ..Sergio PorcherAinda não há avaliações
- 3 - Membrana PlasmáticaDocumento34 páginas3 - Membrana PlasmáticaMarcelo FelipeAinda não há avaliações
- BiologiaDocumento17 páginasBiologiaandreAinda não há avaliações
- Componentes celularesDocumento17 páginasComponentes celularesVera DominguesAinda não há avaliações
- 02 - Citologia e Organização CelularDocumento5 páginas02 - Citologia e Organização CelularMatheus GuimaraesAinda não há avaliações
- Trabalaho Membrana PlasmáticaDocumento16 páginasTrabalaho Membrana PlasmáticaRonny GoisAinda não há avaliações
- Aula6 Membrana PlasmaticaDocumento2 páginasAula6 Membrana PlasmaticaJunior AnotonioAinda não há avaliações
- Membrana celular: estrutura, composição e funçõesDocumento15 páginasMembrana celular: estrutura, composição e funçõesMARIA YASMIN DE SOUSA BERNARDOAinda não há avaliações
- AULA 4 BIOLOGIA CELULAR Estrutura Celular Membrana Plasmática Citoplasma NúcleoDocumento41 páginasAULA 4 BIOLOGIA CELULAR Estrutura Celular Membrana Plasmática Citoplasma Núcleocaroldsll14Ainda não há avaliações
- Aula 3 Estrutua Das MembranasDocumento40 páginasAula 3 Estrutua Das Membranasjachin.trippAinda não há avaliações
- Aula 4 - Biomembranas e OrganelasDocumento56 páginasAula 4 - Biomembranas e OrganelasxisbrjamillyAinda não há avaliações
- VOD - Biologia - Introdução À Citologia e Membrana Plasmática - 2021Documento9 páginasVOD - Biologia - Introdução À Citologia e Membrana Plasmática - 2021Sidney MendesAinda não há avaliações
- Palestra Ricardo OliveiraDocumento77 páginasPalestra Ricardo Oliveiraafonsoedson727Ainda não há avaliações
- Citologia - Membrana Plasmatica 2013 Composicao e AnexosDocumento49 páginasCitologia - Membrana Plasmatica 2013 Composicao e AnexosBeboc Deagonal UlfricoAinda não há avaliações
- Conceitos Gerais das Membranas CelularesDocumento8 páginasConceitos Gerais das Membranas CelularesDaniela GondimAinda não há avaliações
- Funções da membrana plasmáticaDocumento5 páginasFunções da membrana plasmáticaLoretoAinda não há avaliações
- Guia de Estudos Da Unidade 2 - Citologia e EmbriologiaDocumento13 páginasGuia de Estudos Da Unidade 2 - Citologia e EmbriologiaPastoral Familiar Nossa Senhora do CarmoAinda não há avaliações
- Biologia IiDocumento6 páginasBiologia IieduzinhoyAinda não há avaliações
- Biomembranas e transportes celularesDocumento47 páginasBiomembranas e transportes celularesAlex OjimaAinda não há avaliações
- Atividade 3Documento7 páginasAtividade 3Priscila VidalAinda não há avaliações
- 4º Relatório de Biologia CelularDocumento11 páginas4º Relatório de Biologia CelularalinemsantanaAinda não há avaliações
- Trabalho de FisiologiaDocumento14 páginasTrabalho de FisiologiaAlegre BalançaAinda não há avaliações
- Eb Sem 9Documento153 páginasEb Sem 9mih.carvalho.maAinda não há avaliações
- Estrutura Celular RESUMODocumento2 páginasEstrutura Celular RESUMOMariana RodriguesAinda não há avaliações
- Mã - Dulo 9 - Organelas Citoplasmã - Ticas - 230906 - 093446Documento8 páginasMã - Dulo 9 - Organelas Citoplasmã - Ticas - 230906 - 093446Ana Júlia GonçalvesAinda não há avaliações
- Anatomia PatológicaDocumento168 páginasAnatomia PatológicaThayanie Oliveira Pereira100% (1)
- SanarDocumento21 páginasSanarNatã PortugalAinda não há avaliações
- Membrana PlasmáticaDocumento44 páginasMembrana PlasmáticaThaynara TolentinoAinda não há avaliações
- Membrana CelularDocumento5 páginasMembrana CelularMarcelo ColodetiAinda não há avaliações
- Células e membrana plasmáticaDocumento377 páginasCélulas e membrana plasmáticabeatrizAinda não há avaliações
- Membrana Plasmatica 1 AnosDocumento16 páginasMembrana Plasmatica 1 AnosAna SilvaAinda não há avaliações
- Aula 2 - Membrana Celular e Especializações de MembranaDocumento47 páginasAula 2 - Membrana Celular e Especializações de MembranaMateus Ciro DominguêsAinda não há avaliações
- Membrana CelularDocumento3 páginasMembrana CelularRodrigo KerberAinda não há avaliações
- BiomembranasDocumento15 páginasBiomembranasmati cAinda não há avaliações
- FICHA ESTUDO - CITOLOGIA 1º AnoDocumento3 páginasFICHA ESTUDO - CITOLOGIA 1º AnoThássio MouraAinda não há avaliações
- BACTÉRIAS - GRAM+ e GRAMDocumento18 páginasBACTÉRIAS - GRAM+ e GRAMJoanna FlorAinda não há avaliações
- Tipos de Células e Membrana PlasmáticaDocumento8 páginasTipos de Células e Membrana PlasmáticaEduardo SouzaAinda não há avaliações
- MI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaDocumento4 páginasMI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaEriczinAinda não há avaliações
- Membrana Plasmática: Estrutura, Funções e TransporteDocumento5 páginasMembrana Plasmática: Estrutura, Funções e TransporteMoises FerroAinda não há avaliações
- BiologiaDocumento4 páginasBiologiaGABRIELY FERREIRA KICOT LIMAAinda não há avaliações
- Membrana celular: funções e estruturaDocumento20 páginasMembrana celular: funções e estruturaMario Maganizo JuniorAinda não há avaliações
- AULA 3 BIOLOGIA CELULAR - Membranas Biologicas-1Documento81 páginasAULA 3 BIOLOGIA CELULAR - Membranas Biologicas-1Levi CamoesAinda não há avaliações
- Células eucariontes: estrutura e componentes da membrana plasmáticaDocumento18 páginasCélulas eucariontes: estrutura e componentes da membrana plasmáticaLucas NevesAinda não há avaliações
- Aula Membrama P e Parede CDocumento33 páginasAula Membrama P e Parede CGeorge Da JosinaAinda não há avaliações
- Aula 03 Membrana CelularDocumento77 páginasAula 03 Membrana CelularEmanuelleAinda não há avaliações
- Aula 4Documento12 páginasAula 4Aniceto RosarioAinda não há avaliações
- Yan Mesko Da Silva - Atividade 6 - 1ºEM AnoDocumento3 páginasYan Mesko Da Silva - Atividade 6 - 1ºEM AnoYan Mesko05Ainda não há avaliações
- Membranas CelularesDocumento8 páginasMembranas CelularesMarcela BrasilAinda não há avaliações
- Mosaico FluidoDocumento7 páginasMosaico FluidoSuzana Francisco Wesley GarciaAinda não há avaliações
- Teste de Conhecimento CitologiaDocumento6 páginasTeste de Conhecimento CitologiaMANIFESTO PORTO REALAinda não há avaliações
- Membrana celular e permeabilidadeDocumento33 páginasMembrana celular e permeabilidadehendrya08Ainda não há avaliações
- Estrutura da membrana21Documento6 páginasEstrutura da membrana21Monica AlexandreAinda não há avaliações
- Células e suas estruturasDocumento16 páginasCélulas e suas estruturasLucas NevesAinda não há avaliações
- Aula 2 - Biologia - CitologiaDocumento16 páginasAula 2 - Biologia - CitologiaEllyzamaAinda não há avaliações
- Membrana Plasmática - Função - Permeabilidade Da MembranaDocumento14 páginasMembrana Plasmática - Função - Permeabilidade Da MembranaHigor BarrosAinda não há avaliações
- Teste de Dna Eprojeto Genoma - 211025 - 212456Documento17 páginasTeste de Dna Eprojeto Genoma - 211025 - 212456pedro victorAinda não há avaliações
- O Que RevisarDocumento1 páginaO Que RevisarHigor BarrosAinda não há avaliações
- Natureza e Estrutura Do Material Genético - BM 2007Documento62 páginasNatureza e Estrutura Do Material Genético - BM 2007Alexandra SalvadoAinda não há avaliações
- Exercicios Bioquimica DPDocumento8 páginasExercicios Bioquimica DPjuliliossi100% (1)
- Oficina Da FotossinteseDocumento9 páginasOficina Da FotossinteseRodrigo MolinaAinda não há avaliações
- Introdução Ao Metabolismo e BioenergéticaDocumento15 páginasIntrodução Ao Metabolismo e BioenergéticaMonaliza MartinsAinda não há avaliações
- FAMERP 2018 - ProvinhaDocumento26 páginasFAMERP 2018 - ProvinhaMariana B AsseffAinda não há avaliações
- Janine Machado Nóbrega e Izabel Cristina Rodrigues Da SilvaDocumento15 páginasJanine Machado Nóbrega e Izabel Cristina Rodrigues Da SilvaIzabel SilvaAinda não há avaliações
- Proteínas: estrutura, propriedades e migração em campo elétricoDocumento3 páginasProteínas: estrutura, propriedades e migração em campo elétricoLuan SantosAinda não há avaliações
- Organização celular: resolução de exercícios sobre o núcleoDocumento12 páginasOrganização celular: resolução de exercícios sobre o núcleoIssufo SumalgyAinda não há avaliações
- OrganelasDocumento28 páginasOrganelasPatricia OliveiraAinda não há avaliações
- Roteiro de Aula Prática - "Cebola"Documento3 páginasRoteiro de Aula Prática - "Cebola"Gabriel teixeiraAinda não há avaliações
- 2aula Aminoacidos (Modo de Compatibilidade)Documento38 páginas2aula Aminoacidos (Modo de Compatibilidade)Elizael De Jesus GonçalvesAinda não há avaliações
- Complexo de Golgi: estrutura e funçõesDocumento5 páginasComplexo de Golgi: estrutura e funçõesForwomenAinda não há avaliações
- Avaliação de Ciências Naturais sobre Nutrição e DigestãoDocumento4 páginasAvaliação de Ciências Naturais sobre Nutrição e DigestãoMarta CharnecaAinda não há avaliações
- Viebahn Haensler2021tradDocumento15 páginasViebahn Haensler2021tradFabio Guedes MoreiraAinda não há avaliações
- Dicionário de Genética e GATTACA - Experiência GenéticaDocumento3 páginasDicionário de Genética e GATTACA - Experiência GenéticaRodrigoAinda não há avaliações
- Componentes orgânicos e inorgânicos das célulasDocumento60 páginasComponentes orgânicos e inorgânicos das célulasjoice vianaAinda não há avaliações
- Bionergética e Substratos EnergéticosDocumento57 páginasBionergética e Substratos EnergéticosVivian Tezzelle83% (6)
- Bioquímica ClínicaDocumento3 páginasBioquímica ClínicaNilda SouzaAinda não há avaliações
- Membranas Celulares e TransporteDocumento12 páginasMembranas Celulares e TransporteMacedo GeloAinda não há avaliações
- Bioquímica Aplicada IIDocumento75 páginasBioquímica Aplicada IIjackspolAinda não há avaliações
- Lamina - MeiviewDocumento2 páginasLamina - MeiviewPaulaGiselleCzaikoskiAinda não há avaliações
- Telómeros e envelhecimento celularDocumento8 páginasTelómeros e envelhecimento celularCatarina SáAinda não há avaliações
- Catalogo BioestimuladoresDocumento6 páginasCatalogo BioestimuladoresPaula Guima100% (1)
- Genômica Funcional e Epidemiologia GenéticaDocumento14 páginasGenômica Funcional e Epidemiologia Genéticarack26Ainda não há avaliações
- @vetezoo - BIOQUÍMICA CLÍNICA VETERINÁRIA 1Documento36 páginas@vetezoo - BIOQUÍMICA CLÍNICA VETERINÁRIA 1Maria Raquel SilvaAinda não há avaliações
- Bioquímica - Aula 12 - Metabolismo Dos LipídiosDocumento29 páginasBioquímica - Aula 12 - Metabolismo Dos LipídiosTássio BrunoAinda não há avaliações
- Ponto IsoelétricoDocumento3 páginasPonto IsoelétricoAdriana BiasiAinda não há avaliações
- Apostila Ribossomos 2012Documento2 páginasApostila Ribossomos 2012maenagalvaoAinda não há avaliações
- Biópsia de vilo corial e amniocentese: intervenções no pré-natalDocumento17 páginasBiópsia de vilo corial e amniocentese: intervenções no pré-natalMAISAMSBAinda não há avaliações
- Aula de Bioquimica CelularDocumento46 páginasAula de Bioquimica CelularanaaluumontessAinda não há avaliações
- Equilíbrio iônico: aplicações em química analíticaNo EverandEquilíbrio iônico: aplicações em química analíticaAinda não há avaliações
- Simetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaNo EverandSimetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaAinda não há avaliações
- Química da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaNo EverandQuímica da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaAinda não há avaliações
- Detetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseNo EverandDetetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseAinda não há avaliações
- Para Gostar de Química: Introdução à Química GeralNo EverandPara Gostar de Química: Introdução à Química GeralAinda não há avaliações
- Série Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNo EverandSérie Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNota: 5 de 5 estrelas5/5 (1)
- Problemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)No EverandProblemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)Ainda não há avaliações
- Princípios de química analítica: Abordagem teórica qualitativa e quantitativaNo EverandPrincípios de química analítica: Abordagem teórica qualitativa e quantitativaNota: 5 de 5 estrelas5/5 (1)
- Uso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaNo EverandUso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaAinda não há avaliações
- Ácidos e Bases de Brönsted e Lowry: Uma visão aplicada ao meio ambienteNo EverandÁcidos e Bases de Brönsted e Lowry: Uma visão aplicada ao meio ambienteAinda não há avaliações
- Química na Educação Básica: Ferramentas teóricas e práticasNo EverandQuímica na Educação Básica: Ferramentas teóricas e práticasNota: 4 de 5 estrelas4/5 (1)
- Modelagem Matemática de Processos DiversosNo EverandModelagem Matemática de Processos DiversosNota: 5 de 5 estrelas5/5 (4)
- Uma breve história da química: Da alquimia às ciências moleculares modernasNo EverandUma breve história da química: Da alquimia às ciências moleculares modernasAinda não há avaliações
- Química em tubos de ensaio: uma abordagem para principiantesNo EverandQuímica em tubos de ensaio: uma abordagem para principiantesAinda não há avaliações
- Físico-química de Blendas e Compósitos PoliméricosNo EverandFísico-química de Blendas e Compósitos PoliméricosNota: 1 de 5 estrelas1/5 (1)
- Vigilância Sanitária Nos Estados E MunicípiosNo EverandVigilância Sanitária Nos Estados E MunicípiosAinda não há avaliações
- Série Didática para o Apoio à Formação de Professores de Química – Volume 1 – ÁtomosNo EverandSérie Didática para o Apoio à Formação de Professores de Química – Volume 1 – ÁtomosAinda não há avaliações