Você também pode gostar
- Aula 3 Nutrição, Metabolismo, Crescimento e Reprodução BacterianosDocumento59 páginasAula 3 Nutrição, Metabolismo, Crescimento e Reprodução BacterianosEloisa Elena Cangiani100% (3)
- Revisão de MicrobiologiaDocumento14 páginasRevisão de MicrobiologiaJoão Lucas MonteiroAinda não há avaliações
- Biologia Questões 1-20Documento24 páginasBiologia Questões 1-20Antonio CarlosAinda não há avaliações
- Temática - Ciências da Vida - MicrorganismoNo EverandTemática - Ciências da Vida - MicrorganismoAinda não há avaliações
- Cap 1Documento64 páginasCap 1Ricardo PiresAinda não há avaliações
- Produção amidase E. coliDocumento27 páginasProdução amidase E. coliRita BichoAinda não há avaliações
- Nutrição, Metabolismo, Crescimento e Genética Bacteriana: Profa. Dra. Gislaine A. QuerinoDocumento36 páginasNutrição, Metabolismo, Crescimento e Genética Bacteriana: Profa. Dra. Gislaine A. QuerinoPatricia MendesAinda não há avaliações
- 6 Meses para o Enem - NaturezaDocumento46 páginas6 Meses para o Enem - NaturezaHadassa Rebeca rebecaAinda não há avaliações
- 3 Aplicações - Da - BiotecnologiaDocumento54 páginas3 Aplicações - Da - BiotecnologiaHellen LessaAinda não há avaliações
- 100 Questoes Comentadas BIODocumento35 páginas100 Questoes Comentadas BIOFransuelen XavierAinda não há avaliações
- Comentario Apostila 1 Biologia LandimDocumento223 páginasComentario Apostila 1 Biologia Landimdiogocambe04Ainda não há avaliações
- AD1 Microbiologia - Jonatha EgidioDocumento5 páginasAD1 Microbiologia - Jonatha EgidioMatheus PimentelAinda não há avaliações
- Nutrição e probióticos na recuperação de animais debilitadosDocumento8 páginasNutrição e probióticos na recuperação de animais debilitadosFARMACIA petland ma ESTAGIARIOSAinda não há avaliações
- Lista de PáscoaDocumento68 páginasLista de PáscoaMaria Clara Macedo AraújoAinda não há avaliações
- Técnicas de preservação de culturas microbiológicasDocumento6 páginasTécnicas de preservação de culturas microbiológicasBruna Hellen100% (1)
- Apostila de Biotecnologia PDFDocumento28 páginasApostila de Biotecnologia PDFAmanda OliveiraAinda não há avaliações
- Apostila Sobre Proteção E Prevenção Em EnfermagemNo EverandApostila Sobre Proteção E Prevenção Em EnfermagemAinda não há avaliações
- Engenharia Genética-ApostilaDocumento11 páginasEngenharia Genética-ApostilaRei CinucaAinda não há avaliações
- Cultivo de plantas e reprodução seletivaDocumento15 páginasCultivo de plantas e reprodução seletivaAnabelaAinda não há avaliações
- Apostila de Biologia para Vestibular - Módulo 1Documento74 páginasApostila de Biologia para Vestibular - Módulo 1Douglas_Alexan_2145Ainda não há avaliações
- Revisão Aula 1 - História Da Microbiologia e Estrutura Celular BacterianaDocumento9 páginasRevisão Aula 1 - História Da Microbiologia e Estrutura Celular BacterianaKelly SouzaAinda não há avaliações
- Fátima Florêncio Augosto Hélder Americo José Alvaro Fernando Vasco Acacio Rachide Julieta Francisco CébolaDocumento8 páginasFátima Florêncio Augosto Hélder Americo José Alvaro Fernando Vasco Acacio Rachide Julieta Francisco CébolaGidel Joaquim Bacar HageAinda não há avaliações
- Fisiologia Da Regeneração e Da TransplantaçãoDocumento11 páginasFisiologia Da Regeneração e Da TransplantaçãoEdson Vicente100% (1)
- Prova Módulo 2 - UEMDocumento5 páginasProva Módulo 2 - UEMDiego IgorAinda não há avaliações
- Palestra Genoma MilhoDocumento7 páginasPalestra Genoma MilhoAmanda OliveiraAinda não há avaliações
- Extensivoenem-biologia1-Exercícios Sobre Proteínas, Vitaminas e Ácidos NucleicosDocumento6 páginasExtensivoenem-biologia1-Exercícios Sobre Proteínas, Vitaminas e Ácidos NucleicosMaria Eduarda WietholterAinda não há avaliações
- Provas BiologiaDocumento22 páginasProvas BiologiaTeresa silva0% (1)
- Organelas e estruturas celularesDocumento10 páginasOrganelas e estruturas celularesAmanda CamargoAinda não há avaliações
- Lista de Exercícios - Citologia e BiotecDocumento13 páginasLista de Exercícios - Citologia e BiotecMarcella GamaAinda não há avaliações
- Ciclos e Processos Da Sucessão EcológicaDocumento43 páginasCiclos e Processos Da Sucessão EcológicaEdir Edemir ArioliAinda não há avaliações
- 1 Engenharia Genética e DNA RecombinanteDocumento52 páginas1 Engenharia Genética e DNA RecombinanteHellen LessaAinda não há avaliações
- Bactérias: estrutura, reprodução e importânciaDocumento4 páginasBactérias: estrutura, reprodução e importânciakbzAinda não há avaliações
- Práticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNo EverandPráticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNota: 4 de 5 estrelas4/5 (1)
- Apostila BiotecnologiaDocumento41 páginasApostila BiotecnologiaJoão Lucas de AlmeidaAinda não há avaliações
- Revisão III Bimestre - 9º AnoDocumento4 páginasRevisão III Bimestre - 9º AnoAdriAinda não há avaliações
- Evolução do 'oceloide' dos dinoflageladosDocumento10 páginasEvolução do 'oceloide' dos dinoflageladosMariana LopesAinda não há avaliações
- 9-Cultura de Células AnimaisDocumento5 páginas9-Cultura de Células AnimaisMarcela MoraesAinda não há avaliações
- Vod Biologia Biologia ZeroDocumento10 páginasVod Biologia Biologia ZeroMárcia PortoAinda não há avaliações
- Mitocondrias Slide 101121153559 Phpapp01Documento26 páginasMitocondrias Slide 101121153559 Phpapp01Borges Yasmim100% (1)
- Resumos Biologia 4ºtesteDocumento20 páginasResumos Biologia 4ºtesteRafaela CoelhoAinda não há avaliações
- Dieta sem carboidratos e glicogênio muscularDocumento8 páginasDieta sem carboidratos e glicogênio muscularLorrainne CanutoAinda não há avaliações
- A citologia é um ramo da biologia que aprofunda os estudos das células. A teoria celular se desenvolveu principalmente com o surgimento do microscópio. A Biologia Celular, também chamada de Citologia, é a parte daDocumento6 páginasA citologia é um ramo da biologia que aprofunda os estudos das células. A teoria celular se desenvolveu principalmente com o surgimento do microscópio. A Biologia Celular, também chamada de Citologia, é a parte daCamilasiFreitasAinda não há avaliações
- As principais vias metabólicas bacterianas para a obtenção de ATP, NADPH e 13 precursores essenciaisDocumento67 páginasAs principais vias metabólicas bacterianas para a obtenção de ATP, NADPH e 13 precursores essenciaisVale EuropeuAinda não há avaliações
- O Melhoramento-WPS OfficeDocumento60 páginasO Melhoramento-WPS OfficeSimon Juvencio Jose AugustoAinda não há avaliações
- Cultura in Vitro PDFDocumento6 páginasCultura in Vitro PDFxanapat100% (1)
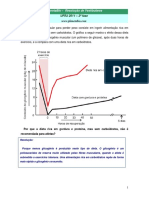
- Curva de Crescimento MicrobianoDocumento9 páginasCurva de Crescimento MicrobianoRita ValleAinda não há avaliações
- 3Documento3 páginas3heltonjapaAinda não há avaliações
- Aula 2 - Classificação e Reprodução Das BactériasDocumento26 páginasAula 2 - Classificação e Reprodução Das BactériasLuís HenriqueAinda não há avaliações
- Trabalho de Biologia Web PDFDocumento21 páginasTrabalho de Biologia Web PDFKelvin Fernando NovidadesAinda não há avaliações
- Reino Monera e seus organismos procariontesDocumento24 páginasReino Monera e seus organismos procariontesKelvin Fernando NovidadesAinda não há avaliações
- Página 1 de 31Documento31 páginasPágina 1 de 31André Sampaio Fontenele de BritoAinda não há avaliações
- Reino MoneraDocumento23 páginasReino MoneraAlessandro ZanardAinda não há avaliações
- Biotecnologia Vegetal: Aplicações e AvançosDocumento6 páginasBiotecnologia Vegetal: Aplicações e AvançosRodolfo Antônio da SilvaAinda não há avaliações
- Cultura de Células e TecidosDocumento40 páginasCultura de Células e TecidosAndré TamAinda não há avaliações
- FT4 - Sintese Proteíca:ciclo CelularDocumento18 páginasFT4 - Sintese Proteíca:ciclo Celularjose eiraAinda não há avaliações
- Clonagem de Plantas IN VITRO PDFDocumento4 páginasClonagem de Plantas IN VITRO PDFSuelen BalbinoAinda não há avaliações
- Resenha Biotecnologia Av2Documento15 páginasResenha Biotecnologia Av2Thay SilvaAinda não há avaliações
- Biossíntese, Funções e Aplicações dos Metabólitos Secundários de PlantasNo EverandBiossíntese, Funções e Aplicações dos Metabólitos Secundários de PlantasAinda não há avaliações
- Pele e AnexosDocumento26 páginasPele e AnexospatriciapppAinda não há avaliações
- Os principais mecanismos de defesa psicológicosDocumento4 páginasOs principais mecanismos de defesa psicológicosFernando Schreiber0% (1)
- Sumário Resumo Inteligencia Emocional LivroDocumento55 páginasSumário Resumo Inteligencia Emocional LivroRamanAinda não há avaliações
- Resumo - Sistema ArticularDocumento3 páginasResumo - Sistema ArticularNajla DiasAinda não há avaliações
- 2-Girl, Forsaken PDFDocumento262 páginas2-Girl, Forsaken PDFMarina Menezes MoratoAinda não há avaliações
- 3 . Aula 04 - EspeciaçãoDocumento24 páginas3 . Aula 04 - EspeciaçãoLéo MaiaAinda não há avaliações
- Guia dos cristais: limpeza, energização e conexãoDocumento37 páginasGuia dos cristais: limpeza, energização e conexãoFabiana Caetano de AssisAinda não há avaliações
- Probabilidades e HeredogramasDocumento15 páginasProbabilidades e HeredogramasAntonio CarlosAinda não há avaliações
- A sexualidade da mulher na terceira idadeDocumento14 páginasA sexualidade da mulher na terceira idadeAugusto SantosAinda não há avaliações
- Cadeia produtiva do algodão no BrasilDocumento30 páginasCadeia produtiva do algodão no BrasilFabio Gonçalves100% (1)
- Resumo - HipercalemiaDocumento7 páginasResumo - HipercalemiaGleycy SubtilAinda não há avaliações
- Importância Dos RiosDocumento24 páginasImportância Dos RiosClaudinha Cris100% (1)
- (BOX 3) Os Melhores Contos Gays - A P WilsonDocumento122 páginas(BOX 3) Os Melhores Contos Gays - A P Wilsonvieiramalu84Ainda não há avaliações
- Fibromialgia: Dor Invisivel (Imagem Corporal e Depressão)Documento12 páginasFibromialgia: Dor Invisivel (Imagem Corporal e Depressão)CDuque100% (12)
- Bioisosterismo: Modificação Molecular RacionalDocumento27 páginasBioisosterismo: Modificação Molecular RacionalElisabeth GomesAinda não há avaliações
- Resumo Wundt Titchener FuncionalismoDocumento13 páginasResumo Wundt Titchener FuncionalismoAmanda PatríciaAinda não há avaliações
- Aula 02 - Medula EspinalDocumento25 páginasAula 02 - Medula EspinalJô BolosAinda não há avaliações
- As pragas da batata e seus danosDocumento11 páginasAs pragas da batata e seus danosBonnerF ApresentadorAinda não há avaliações
- A Terapia de James Hillman: uma psicologia levemente em desacordoDocumento12 páginasA Terapia de James Hillman: uma psicologia levemente em desacordoJD LucasAinda não há avaliações
- Tratamento de feridas com alginato de cálcioDocumento2 páginasTratamento de feridas com alginato de cálcioFelipe ResendeAinda não há avaliações
- Cursos e formações EMAC 2010/2011Documento56 páginasCursos e formações EMAC 2010/2011fcfc02Ainda não há avaliações
- Bem-Estar Na Produção de Alevinos de Tilápia-do-Nilo PDFDocumento54 páginasBem-Estar Na Produção de Alevinos de Tilápia-do-Nilo PDFFábio PaivaAinda não há avaliações
- 3 - Invocacao Da Unificacao Do Chakras (Tashira RenDocumento124 páginas3 - Invocacao Da Unificacao Do Chakras (Tashira Renanon_775332747100% (1)
- Bio Geo 10Documento8 páginasBio Geo 10Pautas TabelasAinda não há avaliações
- Neurose obsessivaDocumento6 páginasNeurose obsessivaDeise Ferreira Dos SantosAinda não há avaliações
- História e nomenclatura dos compostos de coordenaçãoDocumento20 páginasHistória e nomenclatura dos compostos de coordenaçãoCarlos SilvaAinda não há avaliações
- Casqueamento e Ferrageamento em EquinosDocumento53 páginasCasqueamento e Ferrageamento em EquinosConsultor BrêtasAinda não há avaliações
- Kafka Um CruzamentoDocumento2 páginasKafka Um CruzamentoStellaAinda não há avaliações
- O Reformador Do MundoDocumento4 páginasO Reformador Do MundoSilva Kelly GleiceAinda não há avaliações
- 3° - Ano - II - Unidade - CERM - ALUNODocumento7 páginas3° - Ano - II - Unidade - CERM - ALUNOLuiza Antunes Barra MendesAinda não há avaliações