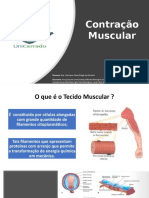
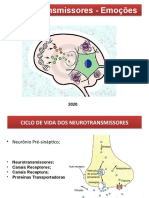
Você também pode gostar
- Aula 2 - Estrutura Da CarneDocumento29 páginasAula 2 - Estrutura Da Carneellen edelynAinda não há avaliações
- Processamento de Carnes - UNIDADE I - Fisiologia Da Carne - 1. Músculo Esquelético - Profa. Dra. Ariana PassosDocumento28 páginasProcessamento de Carnes - UNIDADE I - Fisiologia Da Carne - 1. Músculo Esquelético - Profa. Dra. Ariana Passosantonia.passosAinda não há avaliações
- Resumo Tecido MuscularDocumento4 páginasResumo Tecido MuscularEDUPAinda não há avaliações
- A cannabis nas patologias do sistema nervoso centralNo EverandA cannabis nas patologias do sistema nervoso centralAinda não há avaliações
- FATORESQUEAFETAMAQUALIDADEDACARNEDocumento11 páginasFATORESQUEAFETAMAQUALIDADEDACARNEMarilia C. PantaAinda não há avaliações
- Fisiologia Da ParatireoideDocumento2 páginasFisiologia Da ParatireoideRodrigo DaliaAinda não há avaliações
- Ácidos Graxos de Cadeia Longa Na Saúde e Nutrição PDFDocumento9 páginasÁcidos Graxos de Cadeia Longa Na Saúde e Nutrição PDFWagner CorreiaAinda não há avaliações
- Aula 1 - Propriedades Da MatériaDocumento27 páginasAula 1 - Propriedades Da MatériaIsaias MouraoAinda não há avaliações
- Aula 2 - Estrutura Do MúsculoDocumento55 páginasAula 2 - Estrutura Do MúsculoInayara RebelattoAinda não há avaliações
- 3aula Fisiologia Sistema Nervoso CentralDocumento39 páginas3aula Fisiologia Sistema Nervoso CentralDeborasSouzaAinda não há avaliações
- Estudo Dirigido - FisiologiaDocumento6 páginasEstudo Dirigido - FisiologiaFelipe AssunçãoAinda não há avaliações
- Apostila de Inspeção de Carnes e DerivadosDocumento68 páginasApostila de Inspeção de Carnes e DerivadosGiovana DiogoAinda não há avaliações
- Aspectos Nutricionais e Regulacao Do Metabolismo Na Obesidade e EmagrecimentoDocumento80 páginasAspectos Nutricionais e Regulacao Do Metabolismo Na Obesidade e EmagrecimentoLidiAinda não há avaliações
- Trabalho Contração Muscular FinalDocumento37 páginasTrabalho Contração Muscular FinalNaillyne GalvãoAinda não há avaliações
- Fisiologia Do Baço - TextoDocumento6 páginasFisiologia Do Baço - TextoAleksander MilunovicAinda não há avaliações
- Histologia Humana ResumoDocumento42 páginasHistologia Humana ResumoMedicina I UnilaAinda não há avaliações
- Apresentação 1 - Química 1º EJADocumento9 páginasApresentação 1 - Química 1º EJALucas PontesAinda não há avaliações
- Classes FarmacológicasDocumento10 páginasClasses FarmacológicasBruna Rafaela GomesAinda não há avaliações
- Aula 2 - Sistemas Matria e Estados Fsicos Matria PDFDocumento27 páginasAula 2 - Sistemas Matria e Estados Fsicos Matria PDFGabriel Rodrigues limaAinda não há avaliações
- Resumo - Anatomia e Fisiologia Do EstômagoDocumento5 páginasResumo - Anatomia e Fisiologia Do EstômagoKaiane MaschioAinda não há avaliações
- Exercícios de AgroindustriaDocumento11 páginasExercícios de AgroindustriaCamilly VianaAinda não há avaliações
- Revisao Tecido Muscular e NervosoDocumento14 páginasRevisao Tecido Muscular e NervosoAndrezza CristinaAinda não há avaliações
- Estudo Dirigido EmbriologiaDocumento1 páginaEstudo Dirigido Embriologiavfag100% (1)
- O TECIDO ÓSSEO ResumoDocumento13 páginasO TECIDO ÓSSEO ResumodanieleAinda não há avaliações
- Aula 1 - Biofisica - VeterinariaDocumento26 páginasAula 1 - Biofisica - VeterinariaRodrigo DuarteAinda não há avaliações
- Resumo - Fundamentos Da PatologiaDocumento26 páginasResumo - Fundamentos Da PatologiaMarcoAinda não há avaliações
- Bioquimica GeralDocumento55 páginasBioquimica GeralSamara ErnandesAinda não há avaliações
- Seminário BioquimicaDocumento18 páginasSeminário BioquimicaRoberto CarlosAinda não há avaliações
- Absorção de Nutrientes VegetalDocumento37 páginasAbsorção de Nutrientes VegetalReal AgricolaAinda não há avaliações
- Boas Práticas em Laboratorio - Biologia PDFDocumento270 páginasBoas Práticas em Laboratorio - Biologia PDFAlexandre CasarolliAinda não há avaliações
- Aula LipidiosDocumento24 páginasAula LipidiosJuliana Mendonça100% (1)
- Cortes de CarnesDocumento32 páginasCortes de CarnesJoão Paulo PoltronieriAinda não há avaliações
- Resumo - CitologiaDocumento3 páginasResumo - CitologiaJimmy Lima100% (1)
- Aula 6 - Classes Farmacológicas - MIPS Ou PrincipaisDocumento59 páginasAula 6 - Classes Farmacológicas - MIPS Ou PrincipaisLucas FerreiraAinda não há avaliações
- 03.constituição Da MatériaDocumento34 páginas03.constituição Da MatériawilbletonhankAinda não há avaliações
- 04 08 11 2021 Parasitologia Básica Na EnfermagemDocumento54 páginas04 08 11 2021 Parasitologia Básica Na EnfermagemLótus DouradoAinda não há avaliações
- Regulação Da Secreção Da ACTHDocumento3 páginasRegulação Da Secreção Da ACTHDanilo BernardoAinda não há avaliações
- Beta Oxidação 2017Documento44 páginasBeta Oxidação 2017Maria Eduarda Frota100% (1)
- Aula de Psicofarmacologia Emoções SlidesDocumento37 páginasAula de Psicofarmacologia Emoções SlidesThaline LiviaAinda não há avaliações
- Biofísica Da CirculaçãoDocumento7 páginasBiofísica Da CirculaçãoLuciana CassiaAinda não há avaliações
- Relatorio Embriologia (3 Unidade)Documento3 páginasRelatorio Embriologia (3 Unidade)Jecy SilvaAinda não há avaliações
- 2 Sistema DigestórioDocumento20 páginas2 Sistema DigestórioJaqueline Fekete Coelho100% (1)
- Bromatologia - Slide Carne 18.04.2020Documento8 páginasBromatologia - Slide Carne 18.04.2020Iranildo BarbozaAinda não há avaliações
- Placenta MedicinaDocumento49 páginasPlacenta MedicinaBruno Avelar100% (1)
- Bovinos de Carne - em Vida e em CarcaçaDocumento3 páginasBovinos de Carne - em Vida e em CarcaçaEliana E MindoAinda não há avaliações
- Aula 1 - Homeostase e Organização Funcional Do Sistema NervosoDocumento22 páginasAula 1 - Homeostase e Organização Funcional Do Sistema NervosoElizael GoncalvesAinda não há avaliações
- Química - Aula 1 - Matéria e EnergiaDocumento45 páginasQuímica - Aula 1 - Matéria e Energiamarques07meAinda não há avaliações
- Aula 2 - Estrutura Dos NucleonsDocumento33 páginasAula 2 - Estrutura Dos NucleonsAlessandro Lara100% (1)
- Distúrbios Do Crescimento e Da Diferenciação CelularDocumento17 páginasDistúrbios Do Crescimento e Da Diferenciação CelularWalber FilhoAinda não há avaliações
- Aulaaovivo Ebook Semana 11 2017 PDFDocumento168 páginasAulaaovivo Ebook Semana 11 2017 PDFRafael GhestiAinda não há avaliações
- Avaliação de Rotulagem e Do Perfil de Ácidos Graxos em Suplementos de Ômega 3 Comercializados No BrasilDocumento12 páginasAvaliação de Rotulagem e Do Perfil de Ácidos Graxos em Suplementos de Ômega 3 Comercializados No BrasilLudmila MalveziAinda não há avaliações
- Fisiologia Animal ComparadaDocumento238 páginasFisiologia Animal ComparadaAlexandre OliveiraAinda não há avaliações
- Introdução Ao Estudo Da QuímicaDocumento23 páginasIntrodução Ao Estudo Da QuímicaTais SilvaAinda não há avaliações
- Lista Exercícios ProteínasDocumento1 páginaLista Exercícios ProteínasAna Clara SilvaAinda não há avaliações
- Química Orgânica Teórica 3 Ed.Documento69 páginasQuímica Orgânica Teórica 3 Ed.huitamineiro100% (4)
- Estudo Dirigido-Sistema Genital Masculino.Documento5 páginasEstudo Dirigido-Sistema Genital Masculino.Cristina MeloAinda não há avaliações
- A Importância de Comer Peixes - Pesca em FamíliaDocumento20 páginasA Importância de Comer Peixes - Pesca em FamíliaYana KarinaAinda não há avaliações
- NR 10Documento88 páginasNR 10Leal Safety100% (2)
- Apresentação Do PowerPoint - RSDATA - Cartilha Resumida de Envio Ao EsocialDocumento36 páginasApresentação Do PowerPoint - RSDATA - Cartilha Resumida de Envio Ao EsocialGabriel CamposAinda não há avaliações
- PORTARIA #211 DE 11 DE ABRIL DE 2019 - Imprensa NacionalDocumento2 páginasPORTARIA #211 DE 11 DE ABRIL DE 2019 - Imprensa NacionalGabriel CamposAinda não há avaliações
- Checklist nr24 PDFDocumento43 páginasChecklist nr24 PDFGabriel CamposAinda não há avaliações
- Diretório de Serviços Analytics Brasil 2018 2019Documento77 páginasDiretório de Serviços Analytics Brasil 2018 2019De'Souza Soluções em Segurança do Trabalho100% (1)
- Ebook Contribuições Previdenciárias e Seus Impactos Pela Gestão de SSTDocumento40 páginasEbook Contribuições Previdenciárias e Seus Impactos Pela Gestão de SSTGabriel Campos100% (2)
- Trabalho em AlturaDocumento4 páginasTrabalho em AlturaGabriel CamposAinda não há avaliações
- Invoice PDFDocumento3 páginasInvoice PDFGabriel CamposAinda não há avaliações
- EXERCICO Avaliação Do Cargo - MoleiroDocumento1 páginaEXERCICO Avaliação Do Cargo - MoleiroGabriel CamposAinda não há avaliações
- Edital - Fundação Hospitalar Getúlio VargasDocumento32 páginasEdital - Fundação Hospitalar Getúlio VargasGabriel CamposAinda não há avaliações
- Edital Aluno Especial 2020.1 PGDREDESDocumento10 páginasEdital Aluno Especial 2020.1 PGDREDESGabriel CamposAinda não há avaliações
- Resumo Placas Proj CartórioDocumento2 páginasResumo Placas Proj CartórioGabriel CamposAinda não há avaliações
- NR 15 - Anexo N.º 11 - Agentes Químicos - TolerânciaDocumento5 páginasNR 15 - Anexo N.º 11 - Agentes Químicos - TolerânciaDebora MastantuonoAinda não há avaliações
- 8 Passos para Ser Perito JudicialDocumento4 páginas8 Passos para Ser Perito JudicialGabriel CamposAinda não há avaliações
- Guia de Orientações para Espaços Confinados - Versão para EdiçãoDocumento4 páginasGuia de Orientações para Espaços Confinados - Versão para EdiçãoTaquimAinda não há avaliações
- Instruções de Colocação Protetor PcaDocumento1 páginaInstruções de Colocação Protetor PcaGabriel CamposAinda não há avaliações
- Treinamento Brigada de Incêndio PDFDocumento1 páginaTreinamento Brigada de Incêndio PDFGabriel CamposAinda não há avaliações
- Laudo Insalubridade Sorvetes GlacialDocumento12 páginasLaudo Insalubridade Sorvetes GlacialFabiola SouzaAinda não há avaliações
- Edital Eng SegDocumento39 páginasEdital Eng SegGabriel CamposAinda não há avaliações
- Tipos de AncoragemDocumento6 páginasTipos de AncoragemGabriel CamposAinda não há avaliações
- Memorial de Adequação TombadorDocumento2 páginasMemorial de Adequação TombadorGabriel CamposAinda não há avaliações
- Instruções de Colocação Protetor PcaDocumento1 páginaInstruções de Colocação Protetor PcaGabriel CamposAinda não há avaliações
- Ordem de Serviço - EPIDocumento1 páginaOrdem de Serviço - EPIGabriel CamposAinda não há avaliações
- Sílica - Manual Do Trabalhador - 2 EdiçãoDocumento64 páginasSílica - Manual Do Trabalhador - 2 EdiçãoHudson LegnarAinda não há avaliações
- Requisitos Basicos Programa de Gestao de Espacos ConfinadosDocumento11 páginasRequisitos Basicos Programa de Gestao de Espacos ConfinadosSandro Morais SchultzAinda não há avaliações
- NBR 14276 - 2006 - Brigada de Incêndio PDFDocumento37 páginasNBR 14276 - 2006 - Brigada de Incêndio PDFMarco Antonio HoinskiAinda não há avaliações
- Atendimento Pré-HospitalarDocumento11 páginasAtendimento Pré-HospitalarGabriel CamposAinda não há avaliações
- NBR 11861 PDFDocumento16 páginasNBR 11861 PDFFábio Lima100% (2)
- Modelo de POP 2019 HospitalDocumento45 páginasModelo de POP 2019 HospitalGabriel Campos100% (1)
- Modelo de Laudo de Inviabilidade Técnica CBM-RSDocumento4 páginasModelo de Laudo de Inviabilidade Técnica CBM-RSGabriel CamposAinda não há avaliações
- Relatorio - Síntese Do Ácido Acetilsalicílico (Aas) - Passei DiretoDocumento10 páginasRelatorio - Síntese Do Ácido Acetilsalicílico (Aas) - Passei DiretoJOANA SCAPINIAinda não há avaliações
- Bombas PDFDocumento17 páginasBombas PDFadrianobrittoAinda não há avaliações
- Alcalinidade PDFDocumento10 páginasAlcalinidade PDFCamila Bortolatto RodriguesAinda não há avaliações
- PQC Uninove PDFDocumento70 páginasPQC Uninove PDFJoão JoãoAinda não há avaliações
- MI YT18-22 BROCHURA Correto para Impressão YANMARDocumento20 páginasMI YT18-22 BROCHURA Correto para Impressão YANMARAlberto0% (2)
- Site Projeto de Vivência 2017.2 Química Geral 3108 2Documento2 páginasSite Projeto de Vivência 2017.2 Química Geral 3108 2Cristian CruzAinda não há avaliações
- Síntese Da Teoria de Transporte de Sedimentos de Van Rijn (1984)Documento27 páginasSíntese Da Teoria de Transporte de Sedimentos de Van Rijn (1984)samuel_barsanelliAinda não há avaliações
- Descrição Técnica Fertilizantes - AtualizadaDocumento18 páginasDescrição Técnica Fertilizantes - AtualizadaMatheus CostaAinda não há avaliações
- André Vieira - Como Testar Meus HormniosDocumento32 páginasAndré Vieira - Como Testar Meus HormniosJoão SantanaAinda não há avaliações
- CLEAN (Bateria Moura)Documento34 páginasCLEAN (Bateria Moura)Adilson LucaAinda não há avaliações
- Cálculos - Químicos Massa - MolecularDocumento14 páginasCálculos - Químicos Massa - MolecularPaulo GouveiaAinda não há avaliações
- Artigo Tecnico Vasos de PressãoDocumento2 páginasArtigo Tecnico Vasos de PressãoLuciano Marcelo OliveiraAinda não há avaliações
- Ficha de Ciencias 5º Ano Agua PDFDocumento5 páginasFicha de Ciencias 5º Ano Agua PDFAna Pires100% (1)
- Aula 3 - Corte e Serragem e Lista de ExercíciosDocumento10 páginasAula 3 - Corte e Serragem e Lista de ExercíciosGiuseppe Ferrari100% (1)
- Dimensionamento de Sistema de Prevenção e Combate A Incêndio de Uma EdificaçãoDocumento7 páginasDimensionamento de Sistema de Prevenção e Combate A Incêndio de Uma EdificaçãoJosé SauloAinda não há avaliações
- 030 - 8316 Valvula SolenoideDocumento2 páginas030 - 8316 Valvula SolenoideWilliam Will Controle WoodwardAinda não há avaliações
- Creatin in ADocumento2 páginasCreatin in APaulo MattosAinda não há avaliações
- Exercício 2.1 - Química QB70DDocumento2 páginasExercício 2.1 - Química QB70Dgame of hard “GAME OF HARD”Ainda não há avaliações
- Apostila de TERMOLOGIA NEMESIO CODAP 23 Julho 2014 PDFDocumento14 páginasApostila de TERMOLOGIA NEMESIO CODAP 23 Julho 2014 PDFDemétrius OliveiraAinda não há avaliações
- Mancais de RolamentoDocumento25 páginasMancais de Rolamentorereluta100% (1)
- IFMA CodóDocumento8 páginasIFMA CodóAdonias CarvalhoAinda não há avaliações
- Fispq BR PT Lubrax Hydra 68Documento9 páginasFispq BR PT Lubrax Hydra 68JoaoAinda não há avaliações
- Relatorio Sintese Da DibenzalacetonaDocumento4 páginasRelatorio Sintese Da DibenzalacetonamauroqmcAinda não há avaliações
- Catalogo Transformador Seco PDFDocumento2 páginasCatalogo Transformador Seco PDFWILKENSRANIEL9861Ainda não há avaliações
- Eologia Geral e UrbanaDocumento20 páginasEologia Geral e UrbanaThiago Almeida de Carvalho100% (1)
- FAU MACKENZIE Conforto Dos Ambientes I (2015)Documento5 páginasFAU MACKENZIE Conforto Dos Ambientes I (2015)Philip BraunsteinAinda não há avaliações
- Livro - Formulas Cosmeticos PDFDocumento60 páginasLivro - Formulas Cosmeticos PDFAnderson IFSP100% (4)
- Aula 3 - Reatores Não IdeaisDocumento98 páginasAula 3 - Reatores Não IdeaisLeticia Vitola PasettoAinda não há avaliações
- Lista 1 - Fen - Tran - VDocumento3 páginasLista 1 - Fen - Tran - VMax NunesAinda não há avaliações