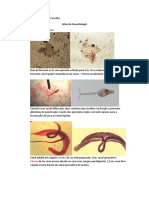
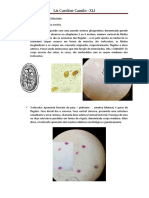
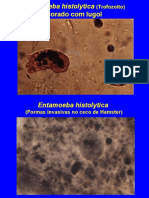
Você também pode gostar
- Microbiologia Médica I: Patógenos e Microbioma HumanoNo EverandMicrobiologia Médica I: Patógenos e Microbioma HumanoAinda não há avaliações
- Atlas ParasitoDocumento16 páginasAtlas ParasitoIsabelleSilvestreAinda não há avaliações
- Ficha de Protozoários 1111Documento7 páginasFicha de Protozoários 1111Efraim Nobre SoaresAinda não há avaliações
- Reino Protista: Doença de ChagasDocumento40 páginasReino Protista: Doença de ChagasFabiane Borges Martins100% (1)
- Patogenia Da Infecção Bacteriana e Resistê Ncia Do Hospedeiro À InfecçãoDocumento27 páginasPatogenia Da Infecção Bacteriana e Resistê Ncia Do Hospedeiro À Infecçãomandy_diiAinda não há avaliações
- Apostila de Imunologia (Pratica)Documento100 páginasApostila de Imunologia (Pratica)yurico122Ainda não há avaliações
- Melo EcologiaDocumento490 páginasMelo Ecologiaceu ceuAinda não há avaliações
- Ação Do Extrato de Própolis Na LeishmanioseDocumento11 páginasAção Do Extrato de Própolis Na LeishmanioseDany VianaAinda não há avaliações
- Artigo 1 Leishmaniose Resposta Imune e Mecanismos Antioxidantes de Escape.p65Documento10 páginasArtigo 1 Leishmaniose Resposta Imune e Mecanismos Antioxidantes de Escape.p65Júnior PintoAinda não há avaliações
- Capitulo 12 A Interacao Do Protozoario Leishmania Com Seus Insetos VetoresDocumento45 páginasCapitulo 12 A Interacao Do Protozoario Leishmania Com Seus Insetos VetoresGabriel LopesAinda não há avaliações
- PBL LeishmanioseDocumento11 páginasPBL LeishmanioseMARIA GRAÇASAinda não há avaliações
- Leishmaniose Visceral Canina RevisatilDocumento8 páginasLeishmaniose Visceral Canina RevisatilFabricio EmídioAinda não há avaliações
- Rodococcus PotroDocumento13 páginasRodococcus Potrohugoh2cnAinda não há avaliações
- Staphylococcus Aureus Meticilina ResistenteDocumento8 páginasStaphylococcus Aureus Meticilina ResistenteSamuelboxAinda não há avaliações
- Vírus e Retrovírus: Contribuição para a Evolução das EspéciesDocumento61 páginasVírus e Retrovírus: Contribuição para a Evolução das EspéciesCristóbal Gutiérrez JiménezAinda não há avaliações
- Apostila Bacteriologia Me Üdica - CortezDocumento121 páginasApostila Bacteriologia Me Üdica - CortezCarolina AlmeidaAinda não há avaliações
- AGD - Estudo Dirigido 31-10-2022Documento3 páginasAGD - Estudo Dirigido 31-10-2022RAFAELA SANTOS FONTANAAinda não há avaliações
- Apostila de Microbiologia IIDocumento83 páginasApostila de Microbiologia IIMarcela RicardoAinda não há avaliações
- Toxoplasmose Revisao PDFDocumento7 páginasToxoplasmose Revisao PDFLEONARDO TÓTOLAAinda não há avaliações
- Imunidade Inata Inflamação Livro Vera CalichDocumento20 páginasImunidade Inata Inflamação Livro Vera CalichAugusto Andrade100% (2)
- Do comensalismo à patogenicidade: Um estudo sobre estafilococos coagulase-negativaNo EverandDo comensalismo à patogenicidade: Um estudo sobre estafilococos coagulase-negativaAinda não há avaliações
- PARASITOLOGIA - Tópico 3 (1)Documento13 páginasPARASITOLOGIA - Tópico 3 (1)katrininovaesAinda não há avaliações
- Artigo Sobre Meningite CriptocócicaDocumento10 páginasArtigo Sobre Meningite CriptocócicaSammuel FreitasAinda não há avaliações
- Pré-Projeto Camilla Malcher Pesset FinalDocumento5 páginasPré-Projeto Camilla Malcher Pesset FinalCamilla Malcher PessetAinda não há avaliações
- Introdução à Parasitologia e seus principais parasitasDocumento50 páginasIntrodução à Parasitologia e seus principais parasitasRafael CedroAinda não há avaliações
- PAIs: Ilhas de patogenicidadeDocumento9 páginasPAIs: Ilhas de patogenicidadeJoao Luiz Macedo0% (1)
- Microbiologia dos FungosDocumento85 páginasMicrobiologia dos FungosFardilha Catarina100% (1)
- Citopato Oncótica - Aula 8Documento67 páginasCitopato Oncótica - Aula 8Rosemari FáveroAinda não há avaliações
- Artigo LeishmaniaDocumento15 páginasArtigo LeishmaniaLuiza PortoAinda não há avaliações
- 3 Forum ParasitologiaDocumento3 páginas3 Forum ParasitologiaLaurindo SurageAinda não há avaliações
- Curso Arena Online 09 Questoes de Biologia Do ENEM Resolvidas e ComentadasDocumento4 páginasCurso Arena Online 09 Questoes de Biologia Do ENEM Resolvidas e Comentadasvanessamariaalt240Ainda não há avaliações
- Recomendações para As Precauções de IsolamentoDocumento47 páginasRecomendações para As Precauções de IsolamentoTiago CunhaAinda não há avaliações
- Artigo MaláriaDocumento12 páginasArtigo MaláriaOctávio Augusto Cruz BritoAinda não há avaliações
- Vírus: causas, transmissão e prevençãoDocumento33 páginasVírus: causas, transmissão e prevençãoEmanuele BazottiAinda não há avaliações
- Introdução à microbiologiaDocumento15 páginasIntrodução à microbiologiaCarynnie KéthileyAinda não há avaliações
- Ruc - ASPECTOS IMUNOLÓGICOS DAS LEISHMANIOSES DERMOTRÓPICAS E VISCEROTRÓPICASDocumento14 páginasRuc - ASPECTOS IMUNOLÓGICOS DAS LEISHMANIOSES DERMOTRÓPICAS E VISCEROTRÓPICASFabio SouzaAinda não há avaliações
- Doenças Casusadas Por FungospdfDocumento17 páginasDoenças Casusadas Por Fungospdfsaul.verasAinda não há avaliações
- Lista de Exercícios (Bacterias e ProtozoáriosDocumento4 páginasLista de Exercícios (Bacterias e ProtozoáriosÁllan David De Araújo SilvaAinda não há avaliações
- Reino MoneraDocumento38 páginasReino Moneragabrieleamorim895Ainda não há avaliações
- Vacina de DNA: aspectos gerais e aplicações na medicina humana e veterináriaDocumento19 páginasVacina de DNA: aspectos gerais e aplicações na medicina humana e veterináriaThiago De Assis CardosoAinda não há avaliações
- Exerci Cio SDocumento18 páginasExerci Cio SSusana SantosAinda não há avaliações
- PARASITOLOGIA - Tópico 4Documento12 páginasPARASITOLOGIA - Tópico 4katrininovaesAinda não há avaliações
- Efetores na interação planta-patógenosDocumento40 páginasEfetores na interação planta-patógenosamandanacional1339Ainda não há avaliações
- Reino Monera PDFDocumento5 páginasReino Monera PDFLuciano JuniorAinda não há avaliações
- Fungos de interesse médico: classificação, patogenia e propriedadesDocumento18 páginasFungos de interesse médico: classificação, patogenia e propriedadesadrienerosceliAinda não há avaliações
- 4 Unid - 4Documento51 páginas4 Unid - 4giselemiranda800Ainda não há avaliações
- Pratica Resposta ImuneDocumento2 páginasPratica Resposta ImuneThaís Sena100% (1)
- Aula 7 Protozoarios 2Documento48 páginasAula 7 Protozoarios 2Carlos Francisco MacuveleAinda não há avaliações
- Aspectos Clínicos Da Toxoplasmose Como Zoonose.Documento21 páginasAspectos Clínicos Da Toxoplasmose Como Zoonose.Pietra Nunes BergerAinda não há avaliações
- trab parasitoDocumento16 páginastrab parasitothais.horanghaeAinda não há avaliações
- Resumo de Nematódeos Intestinais e Lara Migrans VisceralDocumento3 páginasResumo de Nematódeos Intestinais e Lara Migrans Viscerallarissagabriela.limaAinda não há avaliações
- Procura de Um Planeta Sustentável Captura de Tela 2023-06-12 À(s) 21.03.03Documento12 páginasProcura de Um Planeta Sustentável Captura de Tela 2023-06-12 À(s) 21.03.03francinhobhAinda não há avaliações
- Imunologia Do Câncer: Centro Universitário de Brasília Faculadade de Ciências Da SaúdeDocumento31 páginasImunologia Do Câncer: Centro Universitário de Brasília Faculadade de Ciências Da SaúdeMidnight O seisAinda não há avaliações
- Trab Escrito LeishDocumento10 páginasTrab Escrito LeishanaceciliavilacaAinda não há avaliações
- Cap. 07 - Gênero Leishmania - RESUMO PARASITOLOGIADocumento5 páginasCap. 07 - Gênero Leishmania - RESUMO PARASITOLOGIAGustavo SilvaAinda não há avaliações
- Dermatologia No Contexto Da Saúde Única: Dermatofitose Felina de Caráter Zoonótico - Revisão de LiteraturaDocumento13 páginasDermatologia No Contexto Da Saúde Única: Dermatofitose Felina de Caráter Zoonótico - Revisão de LiteraturaSara SantosAinda não há avaliações
- Micro Biologia e ParasitologiaDocumento6 páginasMicro Biologia e ParasitologiaAlciris CorreaAinda não há avaliações
- Parasita IntestinalDocumento19 páginasParasita IntestinalPopinsky JúniorAinda não há avaliações
- Praga Hospitalar: Formicídeos como potenciais transmissores de bactérias em hospitaisNo EverandPraga Hospitalar: Formicídeos como potenciais transmissores de bactérias em hospitaisAinda não há avaliações
- Isso Explicaria Seus Sintomas?: Entendendo os Riscos de Uma Epidemia Oculta da Doença de Lyme no BrasilNo EverandIsso Explicaria Seus Sintomas?: Entendendo os Riscos de Uma Epidemia Oculta da Doença de Lyme no BrasilAinda não há avaliações
- Admin, BJD 569Documento20 páginasAdmin, BJD 569ceu ceuAinda não há avaliações
- Projeto PUB IB 2022-2023Documento6 páginasProjeto PUB IB 2022-2023ceu ceuAinda não há avaliações
- Expressão e purificação de proteínas de P. aeruginosa para estudos de sinalizaçãoDocumento7 páginasExpressão e purificação de proteínas de P. aeruginosa para estudos de sinalizaçãoceu ceuAinda não há avaliações
- Projeto PUB 2022 PESQUISADocumento9 páginasProjeto PUB 2022 PESQUISAceu ceuAinda não há avaliações
- Projeto PUB LCS SBBDocumento10 páginasProjeto PUB LCS SBBceu ceuAinda não há avaliações
- Atlas de ParasitologiaDocumento26 páginasAtlas de ParasitologiaJadson TeglastsonAinda não há avaliações
- Artrópodes: anatomia, fisiologia e taxonomiaDocumento11 páginasArtrópodes: anatomia, fisiologia e taxonomiaAntonio Jose Paulo100% (1)
- CAPITULO AncilostomaDocumento558 páginasCAPITULO AncilostomaWanderson MeloAinda não há avaliações
- Aula 6 Leishmaniose 16Documento36 páginasAula 6 Leishmaniose 16Guilherme MacedoAinda não há avaliações
- Análise crítica do tratamento de cães e gatos com leishmaniose visceralDocumento5 páginasAnálise crítica do tratamento de cães e gatos com leishmaniose visceralTassio QueirozAinda não há avaliações
- Atlas Microbiologia e Parasitologia 0Documento64 páginasAtlas Microbiologia e Parasitologia 0Ana Carla OliveiraAinda não há avaliações
- HMANIADocumento1 páginaHMANIAMurillo MartinAinda não há avaliações
- Centro de Ensino LiteratusDocumento22 páginasCentro de Ensino LiteratusHegon PatrickAinda não há avaliações
- Teste rápido Leishmania Ab ECO Vet para detecção de anticorpos anti-Leishmania em cãesDocumento2 páginasTeste rápido Leishmania Ab ECO Vet para detecção de anticorpos anti-Leishmania em cãesAnimal Care - Espaço VeterinárioAinda não há avaliações
- LeishmanioseDocumento12 páginasLeishmanioseCarlos Eduardo Santos PereiraAinda não há avaliações
- Famema 2024 - Caderno IDocumento12 páginasFamema 2024 - Caderno ImfverissimoAinda não há avaliações
- Protozoários e helmintos em fezesDocumento40 páginasProtozoários e helmintos em fezesHelder Daniel BadianiAinda não há avaliações
- Protozoários: classificação e doenças causadasDocumento52 páginasProtozoários: classificação e doenças causadasBruno Maria OliveiraAinda não há avaliações
- PROTOZOARIOSDocumento11 páginasPROTOZOARIOSDavid Oliveira MarcussoAinda não há avaliações
- Identificação de parasitas e vetores de doençasDocumento9 páginasIdentificação de parasitas e vetores de doençasJoão Guilherme MartiniAinda não há avaliações
- Zoonose causada pela LeishmaniaDocumento4 páginasZoonose causada pela LeishmaniaMariana Sales SouzaAinda não há avaliações
- Banco de imagens de parasitas clínicosDocumento98 páginasBanco de imagens de parasitas clínicoswagneramado100% (1)
- Resumo Leishmaniose Tegumentar AmericanaDocumento4 páginasResumo Leishmaniose Tegumentar AmericanaLeonardo NascimentoAinda não há avaliações
- ParsitoDocumento20 páginasParsitomarceloAinda não há avaliações
- Caracterização histopatológica da leishmaniose visceral canina no DFDocumento116 páginasCaracterização histopatológica da leishmaniose visceral canina no DFDácia ArrudaAinda não há avaliações
- Manual Do MS, Leishmaniose TegumentarDocumento182 páginasManual Do MS, Leishmaniose TegumentarLucianoAinda não há avaliações
- Protozoários e HemoparasitasDocumento144 páginasProtozoários e HemoparasitasBarbara Rauta de AvelarAinda não há avaliações
- LEISHMANIOSE VISCERAL - Uptodate 05.01.20Documento19 páginasLEISHMANIOSE VISCERAL - Uptodate 05.01.20Mariana DóriaAinda não há avaliações
- Cap. 14 - Protozoários FlageladosDocumento10 páginasCap. 14 - Protozoários FlageladosPaulo CarvalhoAinda não há avaliações
- Mentoria Ebserh Febre Maculosa Doenca de Chagas Leishmaniose Leptospirose e MalariaDocumento52 páginasMentoria Ebserh Febre Maculosa Doenca de Chagas Leishmaniose Leptospirose e MalariaCarlos Daniel Palombo ElenterioAinda não há avaliações
- Escopo PCDT Leishmaniosetegumentar EnqueteDocumento21 páginasEscopo PCDT Leishmaniosetegumentar Enquetecsaf.semus.sjrAinda não há avaliações
- Clinica Veterinaria 71Documento116 páginasClinica Veterinaria 71Jackie NunesAinda não há avaliações
- 2021 Revista Humanidades Sertanejas v.6 n.1Documento141 páginas2021 Revista Humanidades Sertanejas v.6 n.1Iago RamonAinda não há avaliações