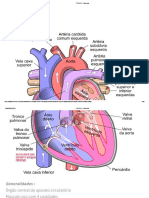
Você também pode gostar
- Divisões Do TelencéfaloDocumento6 páginasDivisões Do TelencéfaloVictor AlencarAinda não há avaliações
- Atividade prática de sistemas tegumentar, locomotor e reprodutorDocumento3 páginasAtividade prática de sistemas tegumentar, locomotor e reprodutorthifesoAinda não há avaliações
- 1 PBDocumento16 páginas1 PBDenilson SantanaAinda não há avaliações
- Projeto de EnsinoDocumento21 páginasProjeto de Ensinoxxi.paulo95Ainda não há avaliações
- Climatogeografia 1aDocumento3 páginasClimatogeografia 1aWailuco AgostinhoAinda não há avaliações
- 36 3 197-204revDocumento8 páginas36 3 197-204revRuan VulcãoAinda não há avaliações
- O Papel e Função Dos Ecossistemas Terrestres e Aquáticos em Moçambique e No - CópiaDocumento4 páginasO Papel e Função Dos Ecossistemas Terrestres e Aquáticos em Moçambique e No - CópiaEliseu Júlio MendesAinda não há avaliações
- Ecossistema (Formativo)Documento3 páginasEcossistema (Formativo)Afonso CanelhoAinda não há avaliações
- Mecanismos de sobrevivência dos coraisDocumento15 páginasMecanismos de sobrevivência dos coraisMiguel RochaAinda não há avaliações
- Algas Nocivas PDFDocumento30 páginasAlgas Nocivas PDFPatricia ChiossiAinda não há avaliações
- EutrifizaçãoDocumento10 páginasEutrifizaçãoRussellAinda não há avaliações
- Macroinvertebrados Bentônicos Como Bioindicadores de Qualidade Ambiental de Corpos Aquáticos Da CaatingaDocumento13 páginasMacroinvertebrados Bentônicos Como Bioindicadores de Qualidade Ambiental de Corpos Aquáticos Da CaatingaEduardoAinda não há avaliações
- Os impactos do aquecimento global na biodiversidade de invertebradosDocumento24 páginasOs impactos do aquecimento global na biodiversidade de invertebradosYuri DantasAinda não há avaliações
- Biologia - CASD - Cadeias e Teias AlimentaresDocumento6 páginasBiologia - CASD - Cadeias e Teias AlimentaresBiologia Apostilas100% (3)
- Ciclos Biogeoquimicos - noPWDocumento56 páginasCiclos Biogeoquimicos - noPWRosberg MotaAinda não há avaliações
- Macroinvertebrados BentônicosDocumento12 páginasMacroinvertebrados BentônicosaderguilhermeAinda não há avaliações
- 2018 - Ecofisiologia - Cap1 e Mudanças ClimáticasDocumento10 páginas2018 - Ecofisiologia - Cap1 e Mudanças ClimáticasknybAinda não há avaliações
- Ficha de Revisão - Biologia 10Documento13 páginasFicha de Revisão - Biologia 10Beatriz SantosAinda não há avaliações
- Priscila Gonçalves Silva - Artigo Indicadores AdemirDocumento16 páginasPriscila Gonçalves Silva - Artigo Indicadores AdemirPriscilaAinda não há avaliações
- Insetos (Insecta Hymenoptera, Lepidoptera e Odonata) e As Mudanças ClimáticasDocumento11 páginasInsetos (Insecta Hymenoptera, Lepidoptera e Odonata) e As Mudanças ClimáticasBreno LoboAinda não há avaliações
- Apostila Rodofíceas Marinhas Bentônicas e As Mudanças ClimáticasDocumento11 páginasApostila Rodofíceas Marinhas Bentônicas e As Mudanças ClimáticasAna Lívia Negrão RibeiroAinda não há avaliações
- Cancian Et Al. 2009Documento6 páginasCancian Et Al. 2009Sandria Ferreira CavassaniAinda não há avaliações
- Red & Peach Illustration World Blood Donor Day FlyerDocumento6 páginasRed & Peach Illustration World Blood Donor Day Flyerliihviviane70Ainda não há avaliações
- BioindicadoresDocumento6 páginasBioindicadoresTessa AltmanAinda não há avaliações
- Lista de exercícios para o ENEMDocumento5 páginasLista de exercícios para o ENEMLucas LkAinda não há avaliações
- Versao Caiola Rumo Ao 20Documento12 páginasVersao Caiola Rumo Ao 20Esm KátyaAinda não há avaliações
- As Plantas e As Mudanças Climáticas - Como Sobreviver em Um Ambiente em Rápida e Constante Transformação - DarwinianasDocumento6 páginasAs Plantas e As Mudanças Climáticas - Como Sobreviver em Um Ambiente em Rápida e Constante Transformação - DarwinianasVictor HegasAinda não há avaliações
- Artigo - Alan Machado SantanaDocumento20 páginasArtigo - Alan Machado SantanaIlca da Graça DionisioAinda não há avaliações
- Mudanças Climáticas e o FitoplânctonDocumento34 páginasMudanças Climáticas e o FitoplânctonagathadeoliveiraAinda não há avaliações
- Fisiologia de bactérias oceânicas profundasDocumento14 páginasFisiologia de bactérias oceânicas profundasEletícia SousaAinda não há avaliações
- Oceanografia BiológicaDocumento33 páginasOceanografia BiológicajoanabrandaoAinda não há avaliações
- 2º TC - Aula 02 - Biodiversidade - Sepam - 2024Documento4 páginas2º TC - Aula 02 - Biodiversidade - Sepam - 2024cardoAinda não há avaliações
- 2º TC - Aula 02 - Biodiversidade - Sepam - 2024Documento4 páginas2º TC - Aula 02 - Biodiversidade - Sepam - 2024Natália HidalgoAinda não há avaliações
- 3 EM 2ºbim - Avaliação 1.Documento4 páginas3 EM 2ºbim - Avaliação 1.Andressa Soares Rezende BarbosaAinda não há avaliações
- Arqueologia I – Análise da fauna pré-históricaDocumento9 páginasArqueologia I – Análise da fauna pré-históricaJúlia DiasAinda não há avaliações
- Apostila Prática QAM 1 Sem 2012Documento26 páginasApostila Prática QAM 1 Sem 2012Ludmila RochaAinda não há avaliações
- Atividade 10 Conservacao Ambientes AquaticosDocumento3 páginasAtividade 10 Conservacao Ambientes AquaticosMarcos PereiraAinda não há avaliações
- Ecologia no ENEMDocumento36 páginasEcologia no ENEMpauloAinda não há avaliações
- Lista de exercícios sobre ciclos biogeoquímicos na UFUDocumento1 páginaLista de exercícios sobre ciclos biogeoquímicos na UFUkarllos eduardo gonzaga amâncioAinda não há avaliações
- As mudanças climáticas e suas influências nas macroalgas marinhas vermelhasDocumento11 páginasAs mudanças climáticas e suas influências nas macroalgas marinhas vermelhasAna Lívia Negrão RibeiroAinda não há avaliações
- Caracterização e importância das macrófitas aquáticas em ambientes límnicosDocumento22 páginasCaracterização e importância das macrófitas aquáticas em ambientes límnicosDerecke SanchesAinda não há avaliações
- Água - Aula 1Documento5 páginasÁgua - Aula 1lucasmanjosAinda não há avaliações
- Pet. Eja 1 Noturno Ok.Documento15 páginasPet. Eja 1 Noturno Ok.TatianeCandida100% (1)
- Ciclos biogeoquímicos do carbono e da águaDocumento1 páginaCiclos biogeoquímicos do carbono e da águaPalomaRodrigues100% (1)
- Barbosa2016 en PTDocumento4 páginasBarbosa2016 en PTGabriel SilvaAinda não há avaliações
- Remoção de nutrientes em esgoto sanitárioDocumento19 páginasRemoção de nutrientes em esgoto sanitárioYasmin MarcosAinda não há avaliações
- Resolução de Questões Abordando Temas Ligados A Ecologia e Meio Ambiente No Enem - DamiaoDocumento18 páginasResolução de Questões Abordando Temas Ligados A Ecologia e Meio Ambiente No Enem - DamiaoDamião OliveiraAinda não há avaliações
- Grupo Experimental Biologia - Dafnias PDFDocumento2 páginasGrupo Experimental Biologia - Dafnias PDFCarolina AroucaAinda não há avaliações
- QA1Documento4 páginasQA1Joana MarquesAinda não há avaliações
- Ae bf10 Qa 1Documento4 páginasAe bf10 Qa 1Glennda Jorge Pinheiro100% (1)
- Fisiologia Dos Animais MergulhadoresDocumento25 páginasFisiologia Dos Animais MergulhadoresSuelen PereiraAinda não há avaliações
- Dia Mundial Dos Oceanos - 4 Ameaças Que Os Colocam em Perigo - National GeographicDocumento9 páginasDia Mundial Dos Oceanos - 4 Ameaças Que Os Colocam em Perigo - National Geographicsamantha088Ainda não há avaliações
- Biologia Marinha - Atividade ComplementarDocumento4 páginasBiologia Marinha - Atividade ComplementarTaís EduardaAinda não há avaliações
- Efeito EstufaDocumento2 páginasEfeito EstufaDávila Rodrigues MacielAinda não há avaliações
- Questões de ecologia sobre desastres ambientais e sucessão ecológicaDocumento1 páginaQuestões de ecologia sobre desastres ambientais e sucessão ecológicaRenato de FreitasAinda não há avaliações
- Ecologia Básica Estudo DirigidoDocumento4 páginasEcologia Básica Estudo DirigidoNoé Da Sua LaiaAinda não há avaliações
- Artigo 3 - Reginaldo SalesDocumento5 páginasArtigo 3 - Reginaldo SalesReginaldo SalesAinda não há avaliações
- Notas Da Geração Verde SciolyDocumento2 páginasNotas Da Geração Verde SciolyScribdTranslationsAinda não há avaliações
- Fisheries Management Eco - 2019 - Barbosa - Relative Importance of Local and Landscape Variables On Fish Assemblages in - En.ptDocumento12 páginasFisheries Management Eco - 2019 - Barbosa - Relative Importance of Local and Landscape Variables On Fish Assemblages in - En.ptVictor YuriAinda não há avaliações
- As aves marinhas como indicadores dos ecossistemas marinhosDocumento17 páginasAs aves marinhas como indicadores dos ecossistemas marinhosJúnior SenseyAinda não há avaliações
- Temática - Ciências da Vida - Estrutura e EcossistemaNo EverandTemática - Ciências da Vida - Estrutura e EcossistemaAinda não há avaliações
- Ambientes AlagadosNo EverandAmbientes AlagadosAinda não há avaliações
- Sistema Nervoso PeriféricoDocumento5 páginasSistema Nervoso PeriféricoAnna Paula SilvaAinda não há avaliações
- Inflamação aguda e crônica: processos, diferenças e implicaçõesDocumento27 páginasInflamação aguda e crônica: processos, diferenças e implicaçõesAmada MagalinAinda não há avaliações
- CONTROLE MOVIMENTO - Sistema Nervoso CentralDocumento109 páginasCONTROLE MOVIMENTO - Sistema Nervoso CentralLucas CazatiAinda não há avaliações
- Timpanismo ruminal em bovino por indigestão vagalDocumento3 páginasTimpanismo ruminal em bovino por indigestão vagalallan felipeAinda não há avaliações
- Hemograma Completo: Valor de ReferênciaDocumento8 páginasHemograma Completo: Valor de Referênciamarcilene vitorianoAinda não há avaliações
- Anatomia e fisiologia dos sistemas do corpo humanoDocumento3 páginasAnatomia e fisiologia dos sistemas do corpo humanoAna Letícia AlcântaraAinda não há avaliações
- Modificações GravídicasDocumento2 páginasModificações GravídicasPâmella RomeroAinda não há avaliações
- Pà S-Graduaã à o Ebook - GlúteosDocumento15 páginasPà S-Graduaã à o Ebook - GlúteosCarlos SilvaAinda não há avaliações
- Invertebrados - Oitavo AnoDocumento87 páginasInvertebrados - Oitavo AnoJonas DiasAinda não há avaliações
- Resumo Sistema DigestórioDocumento5 páginasResumo Sistema DigestórioDiego MenegussiAinda não há avaliações
- Distúrbios Circulatórios Hiperamia e EdemaDocumento6 páginasDistúrbios Circulatórios Hiperamia e EdemaWesley AmaralAinda não há avaliações
- Aula 1 Conceitos Básicos.2023Documento10 páginasAula 1 Conceitos Básicos.2023meucorretorwilAinda não há avaliações
- 3 Deficiencia de G6PDDocumento10 páginas3 Deficiencia de G6PDrichardbiorjAinda não há avaliações
- Sistema ImunológicoDocumento22 páginasSistema ImunológicoMarcella AndionAinda não há avaliações
- AminoácidosDocumento9 páginasAminoácidospelagioAinda não há avaliações
- Apostila Mobilização NeuralDocumento47 páginasApostila Mobilização NeurallineserafimAinda não há avaliações
- TORAX 2 - EvernoteDocumento20 páginasTORAX 2 - EvernoteDerlis OrtizAinda não há avaliações
- Cap. 11 - Sistema Neuroendócrino 1Documento19 páginasCap. 11 - Sistema Neuroendócrino 1Matheus LimaAinda não há avaliações
- Tdah e NeurocienciasDocumento8 páginasTdah e NeurocienciasLuiza LemosAinda não há avaliações
- Equilibrio Ácido Base PneumologiaDocumento27 páginasEquilibrio Ácido Base PneumologiaSilviabeatoAinda não há avaliações
- Atividades físicas e vídeos educativosDocumento14 páginasAtividades físicas e vídeos educativosScheila Brito PereiraAinda não há avaliações
- Questões de Histologia - Tecido MuscularDocumento3 páginasQuestões de Histologia - Tecido MuscularMarcos PauloAinda não há avaliações
- Kinesio TapingDocumento5 páginasKinesio TapingGuto DrumAinda não há avaliações
- ASII Museu & LegendasDocumento86 páginasASII Museu & LegendasGENETICA MEDICAFMDUPAinda não há avaliações
- Resumo Sistema Nervoso CentralDocumento5 páginasResumo Sistema Nervoso CentraltamiossopolianaAinda não há avaliações
- Sintese de AminoacidosDocumento2 páginasSintese de AminoacidosCarlinha WastterAinda não há avaliações
- Anatomia e fisiologia do SNCDocumento17 páginasAnatomia e fisiologia do SNCCarolina Romao100% (1)
- Trabalho de Educação FisicaDocumento3 páginasTrabalho de Educação FisicaNoah Herlander BeccariAinda não há avaliações