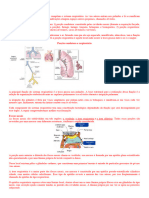
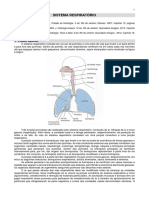
Você também pode gostar
- 9 RespiratDocumento6 páginas9 RespiratElida E KennedyAinda não há avaliações
- Citologia pulmonar e vias respiratóriasDocumento48 páginasCitologia pulmonar e vias respiratóriasCamila SampaioAinda não há avaliações
- Células alveolares e trocas gasosasDocumento36 páginasCélulas alveolares e trocas gasosasVinícius DearmasAinda não há avaliações
- Roteiro Terico Sistema RespiratrioDocumento3 páginasRoteiro Terico Sistema RespiratriohermanomusashiAinda não há avaliações
- Sistema respiratório: funções e constituintesDocumento7 páginasSistema respiratório: funções e constituintesLeonardo SilvaAinda não há avaliações
- ED Fisiologia Respiratória Parte 1 - KandaraDocumento14 páginasED Fisiologia Respiratória Parte 1 - KandaraLouara GomesAinda não há avaliações
- Célula pulmonar: estrutura, funções e importânciaDocumento6 páginasCélula pulmonar: estrutura, funções e importânciaGabriel CarlosAinda não há avaliações
- Sistema respiratório: estrutura e funçãoDocumento4 páginasSistema respiratório: estrutura e funçãoGabriel RosárioAinda não há avaliações
- O sistema respiratório: estrutura, funções e defesaDocumento6 páginasO sistema respiratório: estrutura, funções e defesaMaria EduardaAinda não há avaliações
- Patogenia do Enfisema Pulmonar - Eventos Celulares e MolecularesDocumento4 páginasPatogenia do Enfisema Pulmonar - Eventos Celulares e MolecularesLeoberto Batista Pereira SobrinhoAinda não há avaliações
- DPOC: estrutura pulmonar afetadaDocumento4 páginasDPOC: estrutura pulmonar afetadaReinaldo RiquetAinda não há avaliações
- Histologia do sistema respiratório: estruturas e funçõesDocumento6 páginasHistologia do sistema respiratório: estruturas e funçõesBeatriz GomesAinda não há avaliações
- Sistema ResporatorioDocumento7 páginasSistema ResporatorioWalisson LopesAinda não há avaliações
- 5 Atividade Dos EosinofilosDocumento10 páginas5 Atividade Dos EosinofilosRobertobosco BoscoAinda não há avaliações
- 11-Sistemas RespiratórioDocumento15 páginas11-Sistemas RespiratórioGEYSIANE NUNES BARBOSAAinda não há avaliações
- Fisiologia Respiratória: Funções, Estruturas e Controle NervosoDocumento33 páginasFisiologia Respiratória: Funções, Estruturas e Controle NervosoLouara GomesAinda não há avaliações
- Histologia Do Sistema RespiratórioDocumento10 páginasHistologia Do Sistema RespiratórioSABRINA FURTUNATOAinda não há avaliações
- Aparelho RespiratórioDocumento5 páginasAparelho RespiratórioMaria FernandaAinda não há avaliações
- Vias Aéreas Superiores: Estrutura e Função do NarizDocumento3 páginasVias Aéreas Superiores: Estrutura e Função do NarizStephanie FernandesAinda não há avaliações
- Histologia RespDocumento12 páginasHistologia RespThais LorenaAinda não há avaliações
- Sist RespiratórioDocumento72 páginasSist RespiratórioErika Justo FrigerioAinda não há avaliações
- Resumo Do Sistema RespiratorioDocumento6 páginasResumo Do Sistema RespiratorioSaúde Da MulherAinda não há avaliações
- Sistema Respiratório - ORGANOLOGIADocumento3 páginasSistema Respiratório - ORGANOLOGIALucasBruxelAinda não há avaliações
- Aparelho RespiratorioDocumento2 páginasAparelho RespiratorioFrancisco Bernardes de OliveiraAinda não há avaliações
- HISTOLOGIADocumento7 páginasHISTOLOGIALavinia MedradoAinda não há avaliações
- Livro Texto Unidade IIDocumento39 páginasLivro Texto Unidade IIFrantieskoAinda não há avaliações
- 8ecc - Histologia Do Aparelho RespiratorioDocumento32 páginas8ecc - Histologia Do Aparelho RespiratorioWesleyDimasAinda não há avaliações
- REVISÃO SISTEMA RESPIRATÓRIODocumento18 páginasREVISÃO SISTEMA RESPIRATÓRIOValéria SantosAinda não há avaliações
- Resumo Sistema RespiratórioDocumento6 páginasResumo Sistema RespiratórioLanna RochaAinda não há avaliações
- Sistema Respiratório2Documento24 páginasSistema Respiratório2Alex TauberAinda não há avaliações
- Resumo Do Sistema RespiratorioDocumento6 páginasResumo Do Sistema RespiratorioSaúde Da MulherAinda não há avaliações
- Sistema RespiratórioDocumento10 páginasSistema Respiratóriofisiopaloma9Ainda não há avaliações
- Patogenia do enfisema pulmonarDocumento4 páginasPatogenia do enfisema pulmonarWillian Rafael SampaioAinda não há avaliações
- Sistema RespiratórioDocumento3 páginasSistema RespiratórioRicardo NoronhaAinda não há avaliações
- Histologia do sistema respiratório (40Documento2 páginasHistologia do sistema respiratório (40Maria EduardaAinda não há avaliações
- Embriologia Respiratório e CardiovascularDocumento10 páginasEmbriologia Respiratório e CardiovascularVanessaAinda não há avaliações
- Sistema RespiratórioDocumento9 páginasSistema Respiratórioanabiarosario26Ainda não há avaliações
- Tecido EpitelialDocumento18 páginasTecido EpitelialCarolina Baraky BrederAinda não há avaliações
- Med Resumos - Fisiologia Da RespiraçãoDocumento12 páginasMed Resumos - Fisiologia Da RespiraçãoEsther Beiral100% (3)
- Histologia Do Sistema RespiratórioDocumento6 páginasHistologia Do Sistema RespiratórioThe NerdAinda não há avaliações
- Bronco AspirouDocumento6 páginasBronco AspirouValéria Machado dos SantosAinda não há avaliações
- Relatório Vias AéreasDocumento8 páginasRelatório Vias AéreasVictor NevesAinda não há avaliações
- ALVÉOLOSDocumento4 páginasALVÉOLOSSergio CamaraAinda não há avaliações
- Aula 01 - Fisiologia do sistema respiratório nos animais domésticosDocumento23 páginasAula 01 - Fisiologia do sistema respiratório nos animais domésticosKeyssa MargarizziAinda não há avaliações
- Anatopato RespiratórioDocumento8 páginasAnatopato RespiratórioAna Flávia SilvaAinda não há avaliações
- Histologia RespiratóriaDocumento50 páginasHistologia RespiratóriaBruna BarbetoAinda não há avaliações
- Sistema respiratório: rinite e obstrução brônquicaDocumento21 páginasSistema respiratório: rinite e obstrução brônquicaNaty GiglioAinda não há avaliações
- Sistema respiratório: estrutura e funçõesDocumento13 páginasSistema respiratório: estrutura e funçõesEduardoAinda não há avaliações
- resumo - sist respiratorio (1)Documento15 páginasresumo - sist respiratorio (1)liviacristina.maia09Ainda não há avaliações
- 3 - Sistema RespiratórioDocumento30 páginas3 - Sistema RespiratórioMatheus LanzaAinda não há avaliações
- Nova Síntese SP 13 - Revisão de Sistema Respiratório - Doença Pulmonar Obstrutiva CrônicaDocumento35 páginasNova Síntese SP 13 - Revisão de Sistema Respiratório - Doença Pulmonar Obstrutiva CrônicathaiscavalcanteAinda não há avaliações
- Sistema RespiratórioDocumento2 páginasSistema RespiratórioJuliana VilelaAinda não há avaliações
- Aula 5 - EpitelioDocumento65 páginasAula 5 - Epiteliotuany.sinchettiAinda não há avaliações
- Semiologia do Sistema RespiratórioDocumento21 páginasSemiologia do Sistema RespiratórioHenrique MacetãoAinda não há avaliações
- Sistema respiratório: estrutura e funçãoDocumento54 páginasSistema respiratório: estrutura e funçãoMayara MatosAinda não há avaliações
- Edt Vias Aéreas Superiores Fisiologia PDFDocumento7 páginasEdt Vias Aéreas Superiores Fisiologia PDFHenderson Ferreira100% (1)
- Anatomia do aparelho respiratórioDocumento4 páginasAnatomia do aparelho respiratórioJoabson RibeiroAinda não há avaliações
- Geo 6 Paleomagnetismo ExansaoassoalhooceanicoDocumento16 páginasGeo 6 Paleomagnetismo ExansaoassoalhooceanicoCarlos MarquesAinda não há avaliações
- Geo 13 Rochas MetamorficaDocumento59 páginasGeo 13 Rochas MetamorficaCarlos MarquesAinda não há avaliações
- Geo 4 SismologiaDocumento18 páginasGeo 4 SismologiaCarlos MarquesAinda não há avaliações
- Geo 8 MineraisDocumento36 páginasGeo 8 MineraisCarlos MarquesAinda não há avaliações
- Geo 3 EstruturaInternaTerraDocumento32 páginasGeo 3 EstruturaInternaTerraCarlos MarquesAinda não há avaliações
- Torneio Da Constelação Do Sabre PDFDocumento21 páginasTorneio Da Constelação Do Sabre PDFBeatriz MourãoAinda não há avaliações
- Geo 7 TectonicaPlacasDocumento39 páginasGeo 7 TectonicaPlacasCarlos MarquesAinda não há avaliações
- Geo 10 Rochas IgneasDocumento36 páginasGeo 10 Rochas IgneasCarlos MarquesAinda não há avaliações
- Chave de identificação de plantas brasileirasDocumento87 páginasChave de identificação de plantas brasileirasRicardo OliveiraAinda não há avaliações
- Mighty Blade Manual 3.0 PDFDocumento178 páginasMighty Blade Manual 3.0 PDFequipe rvaAinda não há avaliações
- ArquivototalDocumento93 páginasArquivototalCarlos MarquesAinda não há avaliações
- Baudelaire e A Simbólica Do MalDocumento12 páginasBaudelaire e A Simbólica Do MalMathausSchimidtAinda não há avaliações
- P ApolloniDocumento20 páginasP ApollonirwapolloniAinda não há avaliações
- Morfologia Externa e Organografia de SemDocumento16 páginasMorfologia Externa e Organografia de SemCarlos MarquesAinda não há avaliações
- Identificação dos ossos do esqueleto humanoDocumento2 páginasIdentificação dos ossos do esqueleto humanoThaianne Gadagnoto100% (1)
- Sistema Sensorial O.G.T.A 2019 6 PDFDocumento25 páginasSistema Sensorial O.G.T.A 2019 6 PDFSimone MaiaAinda não há avaliações
- Planimetria e Terminologia Anatômica - Capítulo 3.Documento15 páginasPlanimetria e Terminologia Anatômica - Capítulo 3.Cristine Gondim MaiaAinda não há avaliações
- (Resumo) Imunologia - Aula 2 - Imunidade Inata - Prof. DanielDocumento9 páginas(Resumo) Imunologia - Aula 2 - Imunidade Inata - Prof. DanielMirianAinda não há avaliações
- Anatomia femininaDocumento27 páginasAnatomia femininaSamanta SantarelliAinda não há avaliações
- Zoologia de Vertebrados Laboratório 2Documento7 páginasZoologia de Vertebrados Laboratório 2Adilson JuniorAinda não há avaliações
- Valvas Cardíacas - Anatomia e Relevância Clínica Kenhub 2Documento1 páginaValvas Cardíacas - Anatomia e Relevância Clínica Kenhub 2tuliomariaeduarda7Ainda não há avaliações
- Módulo 3556 Anatomia Aplicada À Epilação DepilaçãoDocumento46 páginasMódulo 3556 Anatomia Aplicada À Epilação Depilaçãombarroso74100% (2)
- 1 - Princípios Gerais Da Técnica CirúrgicaDocumento4 páginas1 - Princípios Gerais Da Técnica Cirúrgicagustavo_dittrichAinda não há avaliações
- Texto Extra Ed Fisica 8o Ano Ens Fundamental II 17032014 1050Documento3 páginasTexto Extra Ed Fisica 8o Ano Ens Fundamental II 17032014 1050Miguel Neto71% (7)
- Anatomia Sistema RespiratórioDocumento53 páginasAnatomia Sistema RespiratórioKelly FariasAinda não há avaliações
- Estudo sobre Neurociências abordando suas principais áreas e conceitosDocumento8 páginasEstudo sobre Neurociências abordando suas principais áreas e conceitosUeslei AndradeAinda não há avaliações
- Prova de Ciências sobre Nutrição e DigestãoDocumento6 páginasProva de Ciências sobre Nutrição e DigestãoDanny BioAinda não há avaliações
- CoagulogramaDocumento39 páginasCoagulogramaverissimoliraAinda não há avaliações
- Introdução à Imunologia: Barreiras, Células e RespostasDocumento55 páginasIntrodução à Imunologia: Barreiras, Células e RespostasLucas LobatoAinda não há avaliações
- Coluna Lombar PDFDocumento93 páginasColuna Lombar PDFKaká Ramos100% (1)
- Lista Exercicio SNADocumento13 páginasLista Exercicio SNADanilo Sousa100% (4)
- Revisão conceitos células tecidosDocumento1 páginaRevisão conceitos células tecidosMaurim Olendino50% (4)
- Artrite ReumatoideDocumento6 páginasArtrite ReumatoideMaria OlíviaAinda não há avaliações
- 1 Esplancnologia PDFDocumento17 páginas1 Esplancnologia PDFJaqueline RodriguesAinda não há avaliações
- Glândulas exócrinas: classificação e estruturaDocumento11 páginasGlândulas exócrinas: classificação e estruturaAngelico Francisco Ângelo BossAinda não há avaliações
- RELATÓRIO DE AULA (Modelo)Documento6 páginasRELATÓRIO DE AULA (Modelo)Andre Rodrigues BorgesAinda não há avaliações
- Sistema UrinárioDocumento12 páginasSistema UrinárioCléber Dos SantosAinda não há avaliações
- Conhecimentos de Medicina Legal e Balística TerminalDocumento29 páginasConhecimentos de Medicina Legal e Balística TerminalMikaira MenezessAinda não há avaliações
- M.T.C. ResumoDocumento1 páginaM.T.C. Resumojailton k100% (2)
- PADRONIZAÇÃO 2 - Arrumação Da Mesa Instrumental, Sinalização, Manuseio Dos Materiais, Manobras Fundamentais e Materiais Especiais - HC I 2021.1Documento16 páginasPADRONIZAÇÃO 2 - Arrumação Da Mesa Instrumental, Sinalização, Manuseio Dos Materiais, Manobras Fundamentais e Materiais Especiais - HC I 2021.1Maria Clara MarinhoAinda não há avaliações
- Pa Tuan Chin - As 8 Peças do BrocadoDocumento7 páginasPa Tuan Chin - As 8 Peças do Brocadojuliano.clj7836Ainda não há avaliações
- Fisiologia - Roteiro de Estudo.Documento7 páginasFisiologia - Roteiro de Estudo.RafaelGregorioAinda não há avaliações
- Trocas Gasosas AnimaisDocumento11 páginasTrocas Gasosas AnimaisVanda CarmoAinda não há avaliações
- Anamnese completa para avaliação médicaDocumento4 páginasAnamnese completa para avaliação médicamirlenalopesAinda não há avaliações