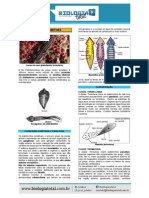
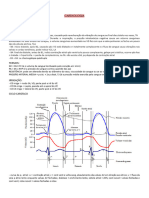
Você também pode gostar
- Introducao Aos Bilateria PlatyhelminthesDocumento46 páginasIntroducao Aos Bilateria Platyhelminthesandrey glauberAinda não há avaliações
- Apostila de Zoologia GeralDocumento30 páginasApostila de Zoologia GeralJulia MaiaAinda não há avaliações
- Filo ArthropodaDocumento19 páginasFilo ArthropodaCaio MaximusAinda não há avaliações
- Provas PPGZOODocumento30 páginasProvas PPGZOOLivia PereiraAinda não há avaliações
- Zoologia de InvertebradosDocumento9 páginasZoologia de InvertebradosSuelen GomesAinda não há avaliações
- Teoria Platelminto NematelmintoDocumento11 páginasTeoria Platelminto NematelmintoMateusMarinhoAinda não há avaliações
- Introducao Aos Eumetazoa Cnidaria CtenophoraDocumento76 páginasIntroducao Aos Eumetazoa Cnidaria Ctenophoraandrey glauberAinda não há avaliações
- PlatelmintosDocumento1 páginaPlatelmintosBianca Gabrielly Oliveira SilvaAinda não há avaliações
- EquinodermosDocumento4 páginasEquinodermoscardoso antonioAinda não há avaliações
- Introdução Ao Filo Arthropoda PDFDocumento21 páginasIntrodução Ao Filo Arthropoda PDFMarlon Casarini MorenoAinda não há avaliações
- RELATÓRIOPRÁTICASEAD - ANATOMIA HUMANA Aula 1 e Aula 2Documento11 páginasRELATÓRIOPRÁTICASEAD - ANATOMIA HUMANA Aula 1 e Aula 2rannyAinda não há avaliações
- Os Poríferos Ou EspongiáriosDocumento6 páginasOs Poríferos Ou EspongiáriosFrancisca Leite de AlmeidaAinda não há avaliações
- Análise Comparativa Entre Os Sistemas de Sustentação Corporal Presentes em AnimaisDocumento57 páginasAnálise Comparativa Entre Os Sistemas de Sustentação Corporal Presentes em AnimaisJéssica MeneghiniAinda não há avaliações
- Zoologia Dos Invertebrados II - InsetosDocumento122 páginasZoologia Dos Invertebrados II - Insetospriiskaf0% (1)
- Características Reino AnimalDocumento3 páginasCaracterísticas Reino AnimalCintia CostaAinda não há avaliações
- Reino Animal CDocumento11 páginasReino Animal CEzimira SaulAinda não há avaliações
- IntroduçãoDocumento21 páginasIntroduçãoMiguel Manuel JoãoAinda não há avaliações
- Filo AntropodesDocumento10 páginasFilo AntropodesmarcostjAinda não há avaliações
- 8 - Zoologia (Parte 01)Documento12 páginas8 - Zoologia (Parte 01)simõesAinda não há avaliações
- AP 1 de Invertebrados IIDocumento26 páginasAP 1 de Invertebrados IIAry JuniorAinda não há avaliações
- Apostila de Anatomia BásicaDocumento78 páginasApostila de Anatomia BásicaAdrianaAinda não há avaliações
- Un. 1 Anatomorfofisiologia Tegumentar, Locomotor, Nervoso e CardiorrespiratórioDocumento107 páginasUn. 1 Anatomorfofisiologia Tegumentar, Locomotor, Nervoso e CardiorrespiratórioFranciele Venturini BarbosaAinda não há avaliações
- CnidáriosDocumento16 páginasCnidáriosSuellen Dias0% (1)
- Trabalhos AnelideosDocumento3 páginasTrabalhos AnelideosBruno MoreiraAinda não há avaliações
- Características Gerais Do Reino AnimaliaDocumento7 páginasCaracterísticas Gerais Do Reino Animaliacb_penatrujillo100% (2)
- Aulas de Anatomia TMG 7 IGCS (1) ActuaDocumento356 páginasAulas de Anatomia TMG 7 IGCS (1) ActuarachidonaboranaboraAinda não há avaliações
- Clivagem, Gastrulação, Mesoderme e NeurulaçãoDocumento7 páginasClivagem, Gastrulação, Mesoderme e NeurulaçãoRaf43la 707Ainda não há avaliações
- Introdução A BilateriaDocumento13 páginasIntrodução A BilateriaJeane AlmeidaAinda não há avaliações
- Reino Animalia 1Documento6 páginasReino Animalia 1Mauro Miguel MartinsAinda não há avaliações
- Aula Introdução Aos Annelida - Polychaeta PDFDocumento176 páginasAula Introdução Aos Annelida - Polychaeta PDFAry JuniorAinda não há avaliações
- O Reino AnimaliaDocumento5 páginasO Reino AnimaliaInfospace100% (1)
- Estudo Dirigido 1 - CoboDocumento2 páginasEstudo Dirigido 1 - CoboNicolli BragaAinda não há avaliações
- Polychaetas ?Documento9 páginasPolychaetas ?Ian Vitor MoreiraAinda não há avaliações
- CORDADOSDocumento4 páginasCORDADOSYone TrindadeAinda não há avaliações
- Reino Animal CARACTERISTICASDocumento11 páginasReino Animal CARACTERISTICASYone TrindadeAinda não há avaliações
- Filo NematodaDocumento8 páginasFilo NematodaHyngrid HevilynAinda não há avaliações
- HelmintosDocumento5 páginasHelmintosluisAinda não há avaliações
- PLATELMINTOSDocumento9 páginasPLATELMINTOSGustavo de BarrosAinda não há avaliações
- BiologiaDocumento20 páginasBiologiaSoares Samuel MuchavaAinda não há avaliações
- Manual de Anatomia e Fisiologia HumanaDocumento46 páginasManual de Anatomia e Fisiologia HumanaFernando FinoAinda não há avaliações
- Apostila de Anatomia e Fisiologia HumanaDocumento46 páginasApostila de Anatomia e Fisiologia HumanaAndreia MonteiroAinda não há avaliações
- Prova Mestrado Zoologia UFRJDocumento39 páginasProva Mestrado Zoologia UFRJKenzel Kache VerborgeAinda não há avaliações
- Os AnelídeosDocumento7 páginasOs AnelídeosJessi FigueraAinda não há avaliações
- Resumo Comparativo Entre Poríferos e CnidáriosDocumento21 páginasResumo Comparativo Entre Poríferos e CnidáriosVânia Da Silva FerreiraAinda não há avaliações
- NematódeosDocumento6 páginasNematódeosRenata BarrosoAinda não há avaliações
- Animais AncelomadosDocumento15 páginasAnimais AncelomadosHelton Victor Camizola CamizolaAinda não há avaliações
- Trabalho de Ciências (Oligochaetas)Documento15 páginasTrabalho de Ciências (Oligochaetas)João Pedro BarsottiAinda não há avaliações
- Ficha 4 12a ClasseDocumento6 páginasFicha 4 12a ClassePenalv Fanny MacanzeAinda não há avaliações
- Resumo Do Capítulo 13Documento18 páginasResumo Do Capítulo 13Felipe Viana TeruelAinda não há avaliações
- Artrópodes - BiologiaDocumento14 páginasArtrópodes - Biologiakrochap123Ainda não há avaliações
- HemoleceDocumento72 páginasHemoleceTayssa MarquesAinda não há avaliações
- Estudo Dirigido de Protostomados para AP 2Documento13 páginasEstudo Dirigido de Protostomados para AP 2Andres RoberAinda não há avaliações
- EMBRIOLOGIA 2022Documento30 páginasEMBRIOLOGIA 2022leonardo citadinAinda não há avaliações
- 11033502032012biologia Invertebrado II Aula 3Documento11 páginas11033502032012biologia Invertebrado II Aula 3588rxt45cvAinda não há avaliações
- Artrópodes e EquinodermosDocumento7 páginasArtrópodes e Equinodermoscz2xhttp4kAinda não há avaliações
- Reino Animalia - Prof. AnacletoDocumento40 páginasReino Animalia - Prof. AnacletoHiper JuniorAinda não há avaliações
- Reino AnimaliaDocumento5 páginasReino AnimaliaAires HumbertoAinda não há avaliações
- FolhasDocumento13 páginasFolhastulioetcetc100% (1)
- Calendário - Calendário AVS 2º Bim.Documento1 páginaCalendário - Calendário AVS 2º Bim.tulioetcetcAinda não há avaliações
- Roteiro de Morfologia VegetalDocumento22 páginasRoteiro de Morfologia VegetaltulioetcetcAinda não há avaliações
- HistologiaDocumento8 páginasHistologiatulioetcetcAinda não há avaliações
- Anexo 1 Formulário Analise de Segurança Da Tarefa - AST - ADocumento3 páginasAnexo 1 Formulário Analise de Segurança Da Tarefa - AST - AThais OliveiraAinda não há avaliações
- Resumo Prova 02Documento32 páginasResumo Prova 02Bruno Félix MendesAinda não há avaliações
- Meridianos Do Coração e Intestino Delgado e Suas Funções Na Medicina Tradicional ChinesaDocumento2 páginasMeridianos Do Coração e Intestino Delgado e Suas Funções Na Medicina Tradicional ChinesaFátima ReginaAinda não há avaliações
- São João Batista Maria Vianney - Biografia, Obras, Espiritualidade - Edemir Lopes de Souza PDFDocumento15 páginasSão João Batista Maria Vianney - Biografia, Obras, Espiritualidade - Edemir Lopes de Souza PDFViam prudentiae100% (2)
- Hematopoiese - EritropoieseDocumento22 páginasHematopoiese - EritropoieseThaís AmorimAinda não há avaliações
- Aula 01 Veiculos Cosmeticos PDFDocumento32 páginasAula 01 Veiculos Cosmeticos PDFRoberta Medeiros100% (1)
- Tireoidite AgudaDocumento2 páginasTireoidite Agudacopiasmed semvirusAinda não há avaliações
- Ebook Receitas Nanda Castelo BrancoDocumento8 páginasEbook Receitas Nanda Castelo Brancofe.brandaoAinda não há avaliações
- Curso para CIPATRDocumento98 páginasCurso para CIPATRAdony AmorimAinda não há avaliações
- Caderno FinalDocumento43 páginasCaderno FinalAlineAinda não há avaliações
- Mapa de QuantidadesDocumento15 páginasMapa de QuantidadesDosmito Joao Desma100% (1)
- Dados Da Fatura de ElectricidadeDocumento3 páginasDados Da Fatura de ElectricidadehermeniodropshippingAinda não há avaliações
- Anamnese DenverDocumento5 páginasAnamnese DenverIndira CamposAinda não há avaliações
- Folder Uso Do EPIDocumento2 páginasFolder Uso Do EPIvsouzajunior100% (2)
- Manual de Formação PDFDocumento19 páginasManual de Formação PDFFátima MoreiraAinda não há avaliações
- Resuminho Sistema RenalDocumento6 páginasResuminho Sistema RenalKarine Fraga MachadoAinda não há avaliações
- A Dor Da CrucifiCAÇãoDocumento3 páginasA Dor Da CrucifiCAÇãojoselmarezendeAinda não há avaliações
- STRALIS 490S40T / 490S44T - 4x2Documento2 páginasSTRALIS 490S40T / 490S44T - 4x2Glauber BrasileiroAinda não há avaliações
- Aspectos Fundamentais Da PsicopedagogiaDocumento3 páginasAspectos Fundamentais Da PsicopedagogiaFranciele De Quadros ColombeliAinda não há avaliações
- Enfermagem Da Saúde Da Criança e Do AdolescenteDocumento21 páginasEnfermagem Da Saúde Da Criança e Do Adolescentenivia ferreiraAinda não há avaliações
- Sandplay Jogo de AreiaDocumento7 páginasSandplay Jogo de Areianeila mesquita100% (1)
- Aula 02: INSS (Técnico Do Seguro Social) Direito Previdenciário - 2022 (Pré-Edital) Prof. Rubens MaurícioDocumento113 páginasAula 02: INSS (Técnico Do Seguro Social) Direito Previdenciário - 2022 (Pré-Edital) Prof. Rubens MaurícioIane SousaAinda não há avaliações
- Ext. Seco Crataegus - Nova LiteraturaDocumento1 páginaExt. Seco Crataegus - Nova LiteraturahowesteveAinda não há avaliações
- 80 - Unimed Vitória - Na05 - Unimed Premium Nacional Coletivo Por Adesao AptDocumento243 páginas80 - Unimed Vitória - Na05 - Unimed Premium Nacional Coletivo Por Adesao AptpessoaanaAinda não há avaliações
- Atualização em Antibiograma Como Implantar As Normas BrCAST e EUCASTDocumento46 páginasAtualização em Antibiograma Como Implantar As Normas BrCAST e EUCASTPatricia Oliver Fiuza100% (1)
- Os Jovens de Dom Bosco Nos Tempos AtuaisDocumento11 páginasOs Jovens de Dom Bosco Nos Tempos AtuaisWilkinson SaturninoAinda não há avaliações
- Bqas-Bio Quantum Analysis System 4.8.6.Documento3 páginasBqas-Bio Quantum Analysis System 4.8.6.rexland1Ainda não há avaliações
- Trabalho de Física - Experiência de Eletrização Por ContatoDocumento9 páginasTrabalho de Física - Experiência de Eletrização Por ContatoDaniel CostaAinda não há avaliações
- CARDIOLOGIADocumento17 páginasCARDIOLOGIAMilena CristinaAinda não há avaliações
- Codex AlimentariusDocumento45 páginasCodex AlimentariusPatrícia EstevesAinda não há avaliações