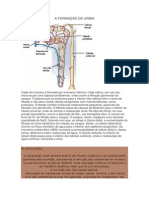
Você também pode gostar
- Resumo de Histologia Renal (Natália Bonfá) 3 PDFDocumento11 páginasResumo de Histologia Renal (Natália Bonfá) 3 PDFTaísa LeirasAinda não há avaliações
- Resumo Fisiologia Renal UfrrjDocumento14 páginasResumo Fisiologia Renal UfrrjM Joana Cesar100% (1)
- Aula 2 - Urinário - RevisadoDocumento73 páginasAula 2 - Urinário - RevisadoEmily RochaAinda não há avaliações
- Intro A Patologia Renal - DirceuDocumento11 páginasIntro A Patologia Renal - DirceuArthur AlvesAinda não há avaliações
- Citologia Do Sistema ExcretorDocumento47 páginasCitologia Do Sistema ExcretorEllen ThamiresAinda não há avaliações
- Patologia Veterinaria - 2 Ed - Santos E Alessi-460-530Documento71 páginasPatologia Veterinaria - 2 Ed - Santos E Alessi-460-530Herick PassosAinda não há avaliações
- Cap 32 - Fisiologia Renal - Berne - ResumoDocumento9 páginasCap 32 - Fisiologia Renal - Berne - ResumoLucas CorrêaAinda não há avaliações
- Sistema Urinário 2008Documento5 páginasSistema Urinário 2008Josi CaldasAinda não há avaliações
- Sistema UrinárioDocumento19 páginasSistema UrinárioPaulo CorreiaAinda não há avaliações
- Abordagem Morfofuncional Do Sistema Urinario PDFDocumento24 páginasAbordagem Morfofuncional Do Sistema Urinario PDFAnderson Soares de AraujoAinda não há avaliações
- TEMA 8 - Fisiologia Do Sistema UrinárioDocumento78 páginasTEMA 8 - Fisiologia Do Sistema UrinárioMaryB.Hbrandt100% (1)
- CAP TULO 6 Fisiologia Excret Ria FormatadoDocumento22 páginasCAP TULO 6 Fisiologia Excret Ria FormatadoEster MontteiroAinda não há avaliações
- Sistema UrinárioDocumento72 páginasSistema UrinárioJulio AlexandreAinda não há avaliações
- Histologia Resumo RimDocumento6 páginasHistologia Resumo RimHitallo BezerraAinda não há avaliações
- Sistema UrinarioDocumento9 páginasSistema UrinarioSABRINA FURTUNATOAinda não há avaliações
- Histologia Do Sistema UrinárioDocumento6 páginasHistologia Do Sistema UrinárioAna Júllia MartinsAinda não há avaliações
- Fisiologia Renal Cap 05Documento13 páginasFisiologia Renal Cap 05Jexica_13Ainda não há avaliações
- Nefrologia - 2020Documento704 páginasNefrologia - 2020Paty1580Ainda não há avaliações
- Fisiologia Renal Resumo 01Documento15 páginasFisiologia Renal Resumo 01Débora Martins100% (1)
- FisiologiaDocumento22 páginasFisiologiaBrunaAinda não há avaliações
- TEMA 06 - Afecções Renais Na InfânciaDocumento21 páginasTEMA 06 - Afecções Renais Na InfânciaAnna Karolinne NascimentoAinda não há avaliações
- Sistema Urinário Histo 2 - Final - Corrigido PDFDocumento10 páginasSistema Urinário Histo 2 - Final - Corrigido PDFGeizibel CoserAinda não há avaliações
- Morfofisiologia Dos NéfronsDocumento10 páginasMorfofisiologia Dos NéfronsBianca AmaralAinda não há avaliações
- (Histologia) Sistema UrinárioDocumento18 páginas(Histologia) Sistema UrinárioSofia VilarAinda não há avaliações
- ResumoDocumento48 páginasResumoViniciusdaSilvaAinda não há avaliações
- Fisio RenalDocumento36 páginasFisio RenalAlmir Fernandes de Carvalho100% (3)
- Resumo Fisiologia RenalDocumento29 páginasResumo Fisiologia RenalHASFTAGAinda não há avaliações
- Aula 08 - Fisiologia Do Sistema UrinárioDocumento61 páginasAula 08 - Fisiologia Do Sistema UrinárioFelipe AssisAinda não há avaliações
- Trabalho HistoDocumento10 páginasTrabalho Histojunosi2004Ainda não há avaliações
- Sistema UrinárioDocumento9 páginasSistema UrinárioKéssia SouzaAinda não há avaliações
- Fisiologia RenalDocumento16 páginasFisiologia RenalAnana AnaAinda não há avaliações
- Nefro - ResumoDocumento283 páginasNefro - ResumoLah FazoloAinda não há avaliações
- T10-11 Aparelho Urinário PDFDocumento7 páginasT10-11 Aparelho Urinário PDFMJTAinda não há avaliações
- Sistema UrinárioDocumento7 páginasSistema UrinárioGuilherme NascimentoAinda não há avaliações
- Resumo Sistema UrinárioDocumento6 páginasResumo Sistema Urináriokriptobebe21Ainda não há avaliações
- Resumo - Fisiologia Renal PDFDocumento24 páginasResumo - Fisiologia Renal PDFLucas De MoraisAinda não há avaliações
- Fisiologia Do Sistema RenalDocumento17 páginasFisiologia Do Sistema RenalFernanda AmaralAinda não há avaliações
- Apostila EdersonDocumento29 páginasApostila EdersonEderson X MarciaAinda não há avaliações
- TRA. Fisiologia RenalDocumento16 páginasTRA. Fisiologia RenalAnaltina VictóriaAinda não há avaliações
- Glândula An A A Digestóri: FígadoDocumento22 páginasGlândula An A A Digestóri: FígadoMaria LetíciaAinda não há avaliações
- Sistema Urinário JunqueiraDocumento4 páginasSistema Urinário JunqueiraRoderley ReisAinda não há avaliações
- Apostila Fisiologia MédicaDocumento57 páginasApostila Fisiologia MédicaTalita RomanichenAinda não há avaliações
- Ficha Informativa - Sistema Urinário - 1Documento9 páginasFicha Informativa - Sistema Urinário - 1Manuela CodêçoAinda não há avaliações
- Fisiologia Dos Rins PDFDocumento8 páginasFisiologia Dos Rins PDFYuri YamasakiAinda não há avaliações
- Filtração Glomerular e Hemodinâmica Renal e Mecanismos de Transporte - CompletoDocumento61 páginasFiltração Glomerular e Hemodinâmica Renal e Mecanismos de Transporte - CompletoFernanda Somera100% (2)
- NefrolitiaseDocumento14 páginasNefrolitiaseMozart WalzAinda não há avaliações
- Tecido MuscularDocumento8 páginasTecido MuscularLAUANE VITORIA CORDEIRO DA FONSECAAinda não há avaliações
- Aps - Corpo Animal Ii - Mirelle LuizaDocumento7 páginasAps - Corpo Animal Ii - Mirelle LuizaMirelle AraújoAinda não há avaliações
- Fisiologia Renal e Formação Da UrinaDocumento13 páginasFisiologia Renal e Formação Da UrinaPery BaccinAinda não há avaliações
- Fisiologia Renal 1Documento17 páginasFisiologia Renal 1Rafaela MendesAinda não há avaliações
- 2 - Sistema Imunologico e LinfaticoDocumento10 páginas2 - Sistema Imunologico e LinfaticoIsabella PimentelAinda não há avaliações
- Histologia - Sistema CirculatórioDocumento57 páginasHistologia - Sistema CirculatórioBruno AvelarAinda não há avaliações
- Sistema UrinárioDocumento11 páginasSistema UrináriomoisesAinda não há avaliações
- Fisiologia Renal - ResumoDocumento24 páginasFisiologia Renal - ResumoRussein C. Franchini100% (3)
- Sistema UrinarioDocumento40 páginasSistema UrinarioMichelle JaraAinda não há avaliações
- A Formação Da UrinaDocumento3 páginasA Formação Da UrinaPedro SantosAinda não há avaliações
- Sistemas Excretores PDFDocumento1 páginaSistemas Excretores PDFNayana JucáAinda não há avaliações
- Sistema Urinario 23EST06 Marcia MeloDocumento12 páginasSistema Urinario 23EST06 Marcia MeloMarcia VianaAinda não há avaliações
- Prova Sistema UrinarioDocumento3 páginasProva Sistema UrinarioCristiano AlvesAinda não há avaliações
- Aula 20Documento21 páginasAula 20Mel RodriguesAinda não há avaliações
- Tese DoutoradoDocumento231 páginasTese DoutoradoRafaellaAinda não há avaliações
- FGV 2022 PC RJ Tecnico Policial de Necropsia ProvaDocumento14 páginasFGV 2022 PC RJ Tecnico Policial de Necropsia ProvaDaniel SilvaAinda não há avaliações
- Nefro IDocumento10 páginasNefro ICinthia TomoeAinda não há avaliações
- Avaliação Ap3Documento9 páginasAvaliação Ap3Leonardo SouzaAinda não há avaliações
- Sistema ExcretorDocumento10 páginasSistema ExcretorMarcelo ColodetiAinda não há avaliações
- Anatomia e Fisiologia Renal - ApostilaDocumento27 páginasAnatomia e Fisiologia Renal - ApostilaLidiane SouzaAinda não há avaliações
- RevisãoDocumento21 páginasRevisãoTércia LemosAinda não há avaliações
- Trabalho Escrito Sistema UrinárioDocumento12 páginasTrabalho Escrito Sistema UrinárioCassia Tavares TavaresAinda não há avaliações
- Urinal Is EsDocumento67 páginasUrinal Is Eselizabeth guimaraes goisAinda não há avaliações
- Sistema UrinárioDocumento27 páginasSistema UrinárioEstokiii D&DAinda não há avaliações
- Embriologia Do Sistema UrinárioDocumento9 páginasEmbriologia Do Sistema UrinárioTamy BeatrizAinda não há avaliações
- Apostila de Farmacologia (3) NovaDocumento89 páginasApostila de Farmacologia (3) NovaDhiego Paixão 2Ainda não há avaliações
- Sistema Urináriodocx - 210512 - 094518Documento14 páginasSistema Urináriodocx - 210512 - 094518Anny LaisAinda não há avaliações
- Ficha de Exercícios (Sistemas Excretor, Reprodutor e Nervoso) - Ii UnidadeDocumento8 páginasFicha de Exercícios (Sistemas Excretor, Reprodutor e Nervoso) - Ii UnidadeJhennifer AzevedoAinda não há avaliações
- Biofísica e A Função Renal.Documento15 páginasBiofísica e A Função Renal.Paulo JoelAinda não há avaliações
- Hormonios e Funcao RenalDocumento35 páginasHormonios e Funcao RenalDanielBCAinda não há avaliações
- respostaPRIMEIRA ATIVIDADE TERCEIRADocumento3 páginasrespostaPRIMEIRA ATIVIDADE TERCEIRATácio MarinhoAinda não há avaliações
- Mapas Mentais AnatomiaDocumento83 páginasMapas Mentais AnatomiaMissão BombeiroAinda não há avaliações
- Ac2 - 1º AnoDocumento6 páginasAc2 - 1º AnoSophia ColaresAinda não há avaliações
- Fisiologia Renal de VanderDocumento26 páginasFisiologia Renal de VanderCarlos Eduardo Avila0% (1)
- Embriologia Do Sistema UrinárioDocumento3 páginasEmbriologia Do Sistema UrinárioRafael Peres100% (1)
- UEM Biologia 2014Documento4 páginasUEM Biologia 2014Sadi Parruque100% (1)
- 12 - Fisiologia Renal - Parte1Documento21 páginas12 - Fisiologia Renal - Parte1GUSTAVO ANDRÉ BARBOSA DE AZEVEDO FILHOAinda não há avaliações