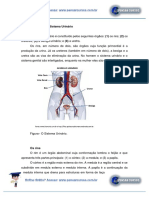
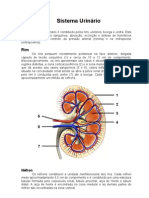
Você também pode gostar
- Patologia Veterinaria - 2 Ed - Santos E Alessi-460-530Documento71 páginasPatologia Veterinaria - 2 Ed - Santos E Alessi-460-530Herick PassosAinda não há avaliações
- Apostila Urinário PDFDocumento14 páginasApostila Urinário PDFEllison Silva AraújoAinda não há avaliações
- Anatomia do Sistema UrinárioDocumento9 páginasAnatomia do Sistema UrinárioVitoria SalesAinda não há avaliações
- Resumo Fisiologia Renal UfrrjDocumento14 páginasResumo Fisiologia Renal UfrrjM Joana Cesar100% (1)
- Histologia Do Sistema UrinárioDocumento6 páginasHistologia Do Sistema UrinárioAna Júllia MartinsAinda não há avaliações
- O sistema urinário: funções, anatomia e fisiologiaDocumento14 páginasO sistema urinário: funções, anatomia e fisiologiaWidek AsfalioAinda não há avaliações
- Sistema urinário: anatomia e fisiologiaDocumento24 páginasSistema urinário: anatomia e fisiologiaAnderson Soares de AraujoAinda não há avaliações
- Cuidados com os rins e doenças renaisDocumento29 páginasCuidados com os rins e doenças renaisEderson X MarciaAinda não há avaliações
- Sistema urinárioDocumento19 páginasSistema urinárioPaulo CorreiaAinda não há avaliações
- Função UrináriaDocumento15 páginasFunção UrináriaTeo SilvaAinda não há avaliações
- Sistema UrinárioDocumento11 páginasSistema UrináriomoisesAinda não há avaliações
- URINARIO LAURA PDDocumento7 páginasURINARIO LAURA PDjofrepereira29Ainda não há avaliações
- O sistema urinário e o néfron: filtração, reabsorção e secreçãoDocumento10 páginasO sistema urinário e o néfron: filtração, reabsorção e secreçãoRayane rodrigues dos santosAinda não há avaliações
- Sistema UrinárioDocumento72 páginasSistema UrinárioJulio AlexandreAinda não há avaliações
- Fisiologa Renal PDFDocumento37 páginasFisiologa Renal PDFGrebyMontalvánRoblesAinda não há avaliações
- 4 Urografia Parte2Documento8 páginas4 Urografia Parte2Rodrigo RibeiroAinda não há avaliações
- TSA29 Appostila Anatomia Fisiologia Farmacologia Renal 2018Documento74 páginasTSA29 Appostila Anatomia Fisiologia Farmacologia Renal 2018Joaquim100% (2)
- Fisiologia Renal Cap 05Documento13 páginasFisiologia Renal Cap 05Jexica_13Ainda não há avaliações
- Fisiologia renal: Funções e estruturas do rimDocumento16 páginasFisiologia renal: Funções e estruturas do rimAnaltina VictóriaAinda não há avaliações
- Intro A Patologia Renal - DirceuDocumento11 páginasIntro A Patologia Renal - DirceuArthur AlvesAinda não há avaliações
- Sistema urinário - anatomia, fisiologia e funçõesDocumento20 páginasSistema urinário - anatomia, fisiologia e funçõesMárcio Stelvio Miranda da SilvaAinda não há avaliações
- 05 - Anatomofisiologia Do Sistema RenalDocumento15 páginas05 - Anatomofisiologia Do Sistema RenalCristiano PereiraAinda não há avaliações
- Fisiologia renal: anatomia, néfron e funções dos segmentos tubularesDocumento48 páginasFisiologia renal: anatomia, néfron e funções dos segmentos tubularesViniciusdaSilvaAinda não há avaliações
- Nefrologia - 2020Documento704 páginasNefrologia - 2020Paty1580Ainda não há avaliações
- Sistema Urinário: Rins, Bexiga e FunçõesDocumento47 páginasSistema Urinário: Rins, Bexiga e FunçõesEllen ThamiresAinda não há avaliações
- Aulas Anatomia e Fisiologia - Parte 5 - S. Urinário e S. ReprodutoresDocumento86 páginasAulas Anatomia e Fisiologia - Parte 5 - S. Urinário e S. ReprodutoresGladisson RamosAinda não há avaliações
- RESUMO - Anatomia 1 - Sistema UrinárioDocumento9 páginasRESUMO - Anatomia 1 - Sistema UrinárioLorena VidreiraAinda não há avaliações
- Fisiologia Renal e Formação Da UrinaDocumento13 páginasFisiologia Renal e Formação Da UrinaPery BaccinAinda não há avaliações
- Anatomia do Sistema UrinárioDocumento18 páginasAnatomia do Sistema UrinárioSofia VilarAinda não há avaliações
- FisiologiaDocumento22 páginasFisiologiaBrunaAinda não há avaliações
- Maria Resumo CaioDocumento15 páginasMaria Resumo CaiovivianferreirapAinda não há avaliações
- 1 Avaliação Laboratorial Do Sistema UrinárioDocumento58 páginas1 Avaliação Laboratorial Do Sistema UrináriorilixgsimululadorAinda não há avaliações
- Sistema Urinário Histo 2 - Final - Corrigido PDFDocumento10 páginasSistema Urinário Histo 2 - Final - Corrigido PDFGeizibel CoserAinda não há avaliações
- Sistema UrinarioDocumento13 páginasSistema Urinariodocemarco21100% (8)
- Morfofisiologia Dos NéfronsDocumento10 páginasMorfofisiologia Dos NéfronsBianca AmaralAinda não há avaliações
- Fisiologia e Patologia Dasdoencas RenaisDocumento26 páginasFisiologia e Patologia Dasdoencas RenaisJuliana Guedes Ferreira LemosAinda não há avaliações
- Resumo Aulas Edson - Sistema UrinárioDocumento9 páginasResumo Aulas Edson - Sistema UrinárioAna Luiza SoutoAinda não há avaliações
- Resumo 1 FrequênciaDocumento106 páginasResumo 1 FrequênciaÉrica CoutinhoAinda não há avaliações
- Filtração Glomerular e Hemodinâmica Renal e Mecanismos de Transporte - CompletoDocumento61 páginasFiltração Glomerular e Hemodinâmica Renal e Mecanismos de Transporte - CompletoFernanda Somera100% (2)
- Anatomia e Fisiologia Do Sistema Genito Urinário - 1Documento7 páginasAnatomia e Fisiologia Do Sistema Genito Urinário - 1marcelodesousameloAinda não há avaliações
- Nutrição e Doenças RenaisDocumento106 páginasNutrição e Doenças RenaisLogain ExcluiAinda não há avaliações
- Aula 2 - Urinário - RevisadoDocumento73 páginasAula 2 - Urinário - RevisadoEmily RochaAinda não há avaliações
- Sistema UrinarioDocumento10 páginasSistema Urinariowallison limaAinda não há avaliações
- Fisiologia Do Sistema RenalDocumento17 páginasFisiologia Do Sistema RenalFernanda AmaralAinda não há avaliações
- Fisio Renal - TortoraDocumento60 páginasFisio Renal - TortoraCamilaAinda não há avaliações
- Fisiologia renal: anatomia e função do néfronDocumento15 páginasFisiologia renal: anatomia e função do néfronDébora Martins100% (1)
- Sistema Urinário: Rins, Néfrons e FunçõesDocumento5 páginasSistema Urinário: Rins, Néfrons e FunçõesJosi CaldasAinda não há avaliações
- Sistema urinárioDocumento9 páginasSistema urinárioManuela CodêçoAinda não há avaliações
- Fisiologia Dos Rins PDFDocumento8 páginasFisiologia Dos Rins PDFYuri YamasakiAinda não há avaliações
- Sistema UrinárioDocumento4 páginasSistema UrinárioMiriam FilipaAinda não há avaliações
- Fisiologia RenalDocumento7 páginasFisiologia RenalNatália OiringAinda não há avaliações
- Apostila Fisiologia MédicaDocumento57 páginasApostila Fisiologia MédicaTalita RomanichenAinda não há avaliações
- Sistema urinário e homeostaseDocumento78 páginasSistema urinário e homeostaseMaryB.Hbrandt100% (1)
- Estudo Do Sistema UrinárioDocumento13 páginasEstudo Do Sistema UrinárioTeo SilvaAinda não há avaliações
- Sistema UrinarioDocumento13 páginasSistema Urinariowallison limaAinda não há avaliações
- Livro Microbiologiaeparasitologia Patologiaeimunologia 2Documento316 páginasLivro Microbiologiaeparasitologia Patologiaeimunologia 2Ester MontteiroAinda não há avaliações
- Cap__tulo_6__Agentes_AntimicrobianosDocumento19 páginasCap__tulo_6__Agentes_AntimicrobianosEster MontteiroAinda não há avaliações
- O Passo-A-passo Para Ter Um Negócio EscalávelDocumento44 páginasO Passo-A-passo Para Ter Um Negócio EscalávelEster MontteiroAinda não há avaliações
- CADERNO_IBFC_181Documento12 páginasCADERNO_IBFC_181Ester MontteiroAinda não há avaliações
- VENTOSA TERAPIADocumento20 páginasVENTOSA TERAPIAEster MontteiroAinda não há avaliações
- EletrotermofototerapiaDocumento21 páginasEletrotermofototerapiaSusan LimaAinda não há avaliações
- aula10-NucleotideoseAcidosNucleicosa530111Documento22 páginasaula10-NucleotideoseAcidosNucleicosa530111Ester MontteiroAinda não há avaliações
- CADERNO_IBFC_107Documento13 páginasCADERNO_IBFC_107Ester MontteiroAinda não há avaliações
- Livro Microbiologiaeparasitologia Patologiaeimunologia 2Documento316 páginasLivro Microbiologiaeparasitologia Patologiaeimunologia 2Ester MontteiroAinda não há avaliações
- eBook Tc Abdome Trifásico Novaead (1)Documento11 páginaseBook Tc Abdome Trifásico Novaead (1)Ester MontteiroAinda não há avaliações
- Posicionamento Radiológico de Crânio (Sala de Raios-X)Documento204 páginasPosicionamento Radiológico de Crânio (Sala de Raios-X)Ester MontteiroAinda não há avaliações
- CAP TULO 2 Fisiologia Geral Excita o El Trica Contra o Muscular e Sistema Nervoso Aut Nomo FormatadoDocumento21 páginasCAP TULO 2 Fisiologia Geral Excita o El Trica Contra o Muscular e Sistema Nervoso Aut Nomo FormatadoEster MontteiroAinda não há avaliações
- CAP TULO 4 Fisiologia Cardiovascular FormatadoDocumento23 páginasCAP TULO 4 Fisiologia Cardiovascular FormatadoEster MontteiroAinda não há avaliações
- CAP TULO 5 Fisiologia Respirat Ria FormatadoDocumento21 páginasCAP TULO 5 Fisiologia Respirat Ria FormatadoEster MontteiroAinda não há avaliações
- Dosagem HDLDocumento7 páginasDosagem HDLAndressaafrancoAinda não há avaliações
- Síntese Artigo - DIF ANAT ENTRE BOVINOS E CÃESDocumento4 páginasSíntese Artigo - DIF ANAT ENTRE BOVINOS E CÃESGlaucia RossattoAinda não há avaliações
- Sistema circulatório e atividade cardíacaDocumento5 páginasSistema circulatório e atividade cardíacaJúlio NascimentoAinda não há avaliações
- IST principais tipos e tratamentosDocumento43 páginasIST principais tipos e tratamentosJanaina MaiaAinda não há avaliações
- Bi Digital O-Ring Test - técnica clínica não invasivaDocumento5 páginasBi Digital O-Ring Test - técnica clínica não invasivaSônia D'AlmeidaAinda não há avaliações
- Descrição CirúrgicaDocumento3 páginasDescrição CirúrgicaIvana Barros100% (4)
- Ginecologia NaturalDocumento90 páginasGinecologia NaturalAline Souza100% (1)
- HipoglicemiantesDocumento2 páginasHipoglicemiantespauloAinda não há avaliações
- Terapia manual e osteopatiaDocumento40 páginasTerapia manual e osteopatiaMarcia RenataAinda não há avaliações
- MC Propriedades e ClassificaçãoDocumento32 páginasMC Propriedades e Classificaçãofabiotcx100% (1)
- Patologias Sobre o Excesso de Proteinas e Carboidratos - MarciaDocumento15 páginasPatologias Sobre o Excesso de Proteinas e Carboidratos - MarciaAlessandro SilvaAinda não há avaliações
- Aula prática sobre sistemas cardiovascular, respiratório e digestórioDocumento1 páginaAula prática sobre sistemas cardiovascular, respiratório e digestórioKerllen MayaraAinda não há avaliações
- Tipos de ReceitaDocumento41 páginasTipos de Receitatiago fortunato94Ainda não há avaliações
- Ebook Guia Introdutório O Poder Dos Óleos EssenciaisDocumento50 páginasEbook Guia Introdutório O Poder Dos Óleos EssenciaisNadine DiasAinda não há avaliações
- Uso de Bicarbonato de Sódio Na Acidose Metabólica PDFDocumento10 páginasUso de Bicarbonato de Sódio Na Acidose Metabólica PDFrasantanaAinda não há avaliações
- Manual Manejo Humanitario de CaesDocumento40 páginasManual Manejo Humanitario de CaeslagartojrAinda não há avaliações
- Paciente idoso internado com suspeita de hepatiteDocumento1 páginaPaciente idoso internado com suspeita de hepatiteMarcos SantanaAinda não há avaliações
- AD2 - Genética 2015.1 Com GabaritoDocumento5 páginasAD2 - Genética 2015.1 Com GabaritoEvenise MotaAinda não há avaliações
- Zumbido na Medicina Tradicional Chinesa: Revisão BibliográficaDocumento16 páginasZumbido na Medicina Tradicional Chinesa: Revisão BibliográficaMarcos RangelAinda não há avaliações
- Modelos LaudosDocumento7 páginasModelos LaudosJailda RodriguesAinda não há avaliações
- 7 Casos Clinicos Antihipertensivos 07-04-2011 0Documento39 páginas7 Casos Clinicos Antihipertensivos 07-04-2011 0Fabricio QuerozAinda não há avaliações
- Teste Psicológico EsaviDocumento20 páginasTeste Psicológico EsaviThay Espindola92% (13)
- Manifestações oculares de doenças sistêmicas: Retinopatia hipertensivaDocumento3 páginasManifestações oculares de doenças sistêmicas: Retinopatia hipertensivasaghacAinda não há avaliações
- Iii ObbDocumento6 páginasIii ObbProfessora CarolAinda não há avaliações
- Exames Laboratoriais - TextoDocumento62 páginasExames Laboratoriais - TextoCatarina ViveirosAinda não há avaliações
- Estudos EndocrinologiaDocumento13 páginasEstudos EndocrinologiaPaula Alves de MouraAinda não há avaliações
- Material Curso de Férias 2022 - Robério PDFDocumento22 páginasMaterial Curso de Férias 2022 - Robério PDFRobério BacelarAinda não há avaliações
- Bula RifocinaDocumento5 páginasBula RifocinaJefferson SilvaAinda não há avaliações
- Biologia AvaliaçãoDocumento4 páginasBiologia AvaliaçãoRenato RibeiroAinda não há avaliações