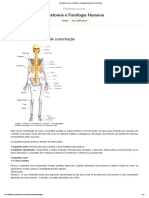
Você também pode gostar
- Tecido MuscularDocumento44 páginasTecido MuscularAna Lucia MarquesAinda não há avaliações
- Arquitetura muscular e contraçãoDocumento120 páginasArquitetura muscular e contraçãoMagno DuarteAinda não há avaliações
- Manual CarnesDocumento10 páginasManual CarnesisabelmartiresAinda não há avaliações
- 13 Tec MuscularDocumento47 páginas13 Tec MuscularmalubastosAinda não há avaliações
- Sistema MuscularDocumento36 páginasSistema MuscularCacalwebAinda não há avaliações
- Procrastinando A Procrastinacao FINALDocumento40 páginasProcrastinando A Procrastinacao FINALestudiosojuliano100% (2)
- Cortes de CarnesDocumento6 páginasCortes de CarnesArlan FscAinda não há avaliações
- HistologiaPL - Tecido MuscularDocumento35 páginasHistologiaPL - Tecido Muscularviolleta gomia100% (1)
- Sistema muscular: Mioblastos e formação das fibras muscularesDocumento43 páginasSistema muscular: Mioblastos e formação das fibras muscularesIvanCostaPassosAinda não há avaliações
- Manual Da UFCD 6565 - Noções Gerais Sobre Células, Imunidade, Tecidos e Órgãos - Sistemas Osteo-Articular e MuscularDocumento50 páginasManual Da UFCD 6565 - Noções Gerais Sobre Células, Imunidade, Tecidos e Órgãos - Sistemas Osteo-Articular e MuscularJoana Rangel100% (5)
- Contração do músculo esqueléticoDocumento37 páginasContração do músculo esqueléticoFERNANDO VINÍCIUS ARAUJO DELMONDESAinda não há avaliações
- Questões Sobre o Tecido MuscularDocumento4 páginasQuestões Sobre o Tecido MuscularEmilly Xavier100% (1)
- Membros e articulaçõesDocumento76 páginasMembros e articulaçõesMaurício Jr.Ainda não há avaliações
- Sistema muscular: estrutura e contraçãoDocumento18 páginasSistema muscular: estrutura e contraçãoGessica Fabiana Cardoso0% (1)
- ARTIGO - Fisiologia Do Músculo EsqueléticoDocumento32 páginasARTIGO - Fisiologia Do Músculo EsqueléticoKeyla SilvaAinda não há avaliações
- Tecido Muscular..Documento8 páginasTecido Muscular..GARDON SINVALDOAinda não há avaliações
- Músculo e DigestãoDocumento19 páginasMúsculo e DigestãomoematimoAinda não há avaliações
- Tecido MuscularDocumento43 páginasTecido Muscular5qgt7tcwxwAinda não há avaliações
- Tecido Muscular EsqueléticoDocumento13 páginasTecido Muscular EsqueléticoMilene Borges MendesAinda não há avaliações
- Fisiologia Músculo Estriado Esquelético 2023Documento36 páginasFisiologia Músculo Estriado Esquelético 2023Fabio Ferreira LorenaAinda não há avaliações
- Tecido MuscularDocumento2 páginasTecido MuscularJULIANA GONÇALVES SAMPAIOAinda não há avaliações
- Tecido MuscularDocumento9 páginasTecido MuscularIsadora RuthAinda não há avaliações
- Fisiologia MuscularDocumento17 páginasFisiologia MuscularMaria Baião BasílioAinda não há avaliações
- Apg 20 - Semestre 2Documento4 páginasApg 20 - Semestre 2Bruna BuenoAinda não há avaliações
- OpenAccess Lorente 9788580393996 08Documento12 páginasOpenAccess Lorente 9788580393996 08Larissa Medicina veterináriaAinda não há avaliações
- Resumo Tecido MuscularDocumento5 páginasResumo Tecido MuscularLaryssa AmorimAinda não há avaliações
- Estrutura e classificação do tecido muscularDocumento10 páginasEstrutura e classificação do tecido muscularThiago RibeiroAinda não há avaliações
- Tecido MuscularDocumento5 páginasTecido MuscularRodrigo SousaAinda não há avaliações
- Tecido MuscularDocumento8 páginasTecido MuscularSIMONE MAIAAinda não há avaliações
- Fisiologia Do Musc EsqueléticoDocumento32 páginasFisiologia Do Musc EsqueléticosalvadoorjrAinda não há avaliações
- Resumo Tecido Muscular Histologia e Fisiologia Da ContraçãoDocumento10 páginasResumo Tecido Muscular Histologia e Fisiologia Da ContraçãoIgor BarbosaAinda não há avaliações
- Fisiologia Muscular (1)Documento9 páginasFisiologia Muscular (1)Ponto XeroxAinda não há avaliações
- Contração muscular: mecanismos e tiposDocumento37 páginasContração muscular: mecanismos e tiposNaillyne GalvãoAinda não há avaliações
- M10P2 - Sistema motor somáticoDocumento8 páginasM10P2 - Sistema motor somáticoana flavia furtado telesAinda não há avaliações
- Sistema Músculo-EsqueléticoDocumento21 páginasSistema Músculo-EsqueléticoiotticarolAinda não há avaliações
- Fisiologia do músculo esqueléticoDocumento57 páginasFisiologia do músculo esqueléticoFábio CastroAinda não há avaliações
- Fechamento Tutoria 12Documento12 páginasFechamento Tutoria 12vanessa da silva caetanoAinda não há avaliações
- TecidosDocumento3 páginasTecidosPedro GuedesAinda não há avaliações
- Tecido MuscularDocumento85 páginasTecido MuscularLarissa MendesAinda não há avaliações
- Tecido MuscularDocumento7 páginasTecido MuscularRebeca SoaresAinda não há avaliações
- TECIDO MUSCULAR - power pointDocumento37 páginasTECIDO MUSCULAR - power pointrjs.mendesAinda não há avaliações
- Histologia Do Tecido Muscular @GusthaDiasDocumento3 páginasHistologia Do Tecido Muscular @GusthaDiasAna Clara FrotaAinda não há avaliações
- Relatório de tutoria sobre dor na coluna e fatores de riscoDocumento6 páginasRelatório de tutoria sobre dor na coluna e fatores de riscoFernanda HerênioAinda não há avaliações
- Tecido MuscularDocumento6 páginasTecido MuscularmayraAinda não há avaliações
- Contração MuscularDocumento13 páginasContração Muscularjonas beneditoAinda não há avaliações
- Classificação e características do tecido muscularDocumento63 páginasClassificação e características do tecido muscularGabriella WeidleAinda não há avaliações
- Unidade Motora e Suas SindromesDocumento39 páginasUnidade Motora e Suas SindromesCibele BragaAinda não há avaliações
- Tecidos Musculares 40Documento3 páginasTecidos Musculares 40Raquel ChequeleiroAinda não há avaliações
- Resumo Tecido Muscular - JunqueiraDocumento4 páginasResumo Tecido Muscular - JunqueiraLucas Thiago BorgesAinda não há avaliações
- Tecidos musculares estriados e lisosDocumento6 páginasTecidos musculares estriados e lisosvanuzaAinda não há avaliações
- Contra CPDFDocumento49 páginasContra CPDFGabriel VianaAinda não há avaliações
- Anatomia General Dos Musculos-1Documento13 páginasAnatomia General Dos Musculos-1martagumbauAinda não há avaliações
- Unid 6Documento26 páginasUnid 6marcelisousadasilvaAinda não há avaliações
- FISIOLOGIA - RESUMO - Contração Do Musculo EsqueléticoDocumento2 páginasFISIOLOGIA - RESUMO - Contração Do Musculo EsqueléticoLara Geovana BezerraAinda não há avaliações
- Prática Viii - Soi IiDocumento12 páginasPrática Viii - Soi IiFabricio Rodrimaur Rodrigues GonçalvesAinda não há avaliações
- Sistema Muscular - FisiologiaDocumento5 páginasSistema Muscular - FisiologiaKauan GabrielAinda não há avaliações
- Tecido MuscularDocumento4 páginasTecido MuscularSara AleixoAinda não há avaliações
- Documento de EstudioDocumento3 páginasDocumento de EstudioDayana DbsAinda não há avaliações
- Apostila Crochetagem 2022Documento14 páginasApostila Crochetagem 2022Tayane TavaresAinda não há avaliações
- Tecidos musculares: características e tiposDocumento3 páginasTecidos musculares: características e tiposGabriela PiresAinda não há avaliações
- ESTRUTURA[1]..Documento7 páginasESTRUTURA[1]..Ozzyel MAinda não há avaliações
- FisiologiaDocumento38 páginasFisiologiaAleida SantosAinda não há avaliações
- Movimento e características do tecido muscularDocumento3 páginasMovimento e características do tecido muscularcedaspy cedaspyAinda não há avaliações
- Apg s8p1Documento3 páginasApg s8p1gabrielAinda não há avaliações
- Resumo Contração MuscularDocumento11 páginasResumo Contração MuscularThiago PontesAinda não há avaliações
- Técnicas cirúrgicas minimamente invasivas para artrodese interfalangeana proximal em equinos, uma comparação das propriedades biomecânicas entre dois procedimentosNo EverandTécnicas cirúrgicas minimamente invasivas para artrodese interfalangeana proximal em equinos, uma comparação das propriedades biomecânicas entre dois procedimentosAinda não há avaliações
- ESF Pirinã G2Documento36 páginasESF Pirinã G2estudiosojulianoAinda não há avaliações
- Vasos - digestórioDocumento5 páginasVasos - digestórioestudiosojulianoAinda não há avaliações
- Psico 3 AvDocumento3 páginasPsico 3 AvestudiosojulianoAinda não há avaliações
- Sistema MuscularDocumento5 páginasSistema MuscularJulia de CarvalhoAinda não há avaliações
- HistologiaDocumento11 páginasHistologiaPedro Sutério100% (1)
- Chaves de Correção - Biologia - Emescam - 1 2023Documento3 páginasChaves de Correção - Biologia - Emescam - 1 2023Ramon Coelho SiqueiraAinda não há avaliações
- Fisiologia Musculo EsqueléticoDocumento14 páginasFisiologia Musculo EsqueléticoRickye BarbosaAinda não há avaliações
- PGA 1a SÉRIE 2BIM PDFDocumento20 páginasPGA 1a SÉRIE 2BIM PDFDaniel Lino50% (2)
- Músculos do corpo humano: principais características e funçõesDocumento14 páginasMúsculos do corpo humano: principais características e funçõesRycher AguiarAinda não há avaliações
- Aparelho Locomotor PDFDocumento61 páginasAparelho Locomotor PDFSabrina FerreiraAinda não há avaliações
- TECIDO MUSCULAR E SEUS COMPONENTESDocumento11 páginasTECIDO MUSCULAR E SEUS COMPONENTESDay MedeirosAinda não há avaliações
- Sistema MuscularDocumento3 páginasSistema MuscularJulio SanchezAinda não há avaliações
- Contração muscular: processos de excitação-contração e tipos de contraçãoDocumento21 páginasContração muscular: processos de excitação-contração e tipos de contraçãoPedro AugustoAinda não há avaliações
- Oe Com Capa 2ºb 2 SérieDocumento21 páginasOe Com Capa 2ºb 2 SérieYouAinda não há avaliações
- Slide Tecido MuscularDocumento12 páginasSlide Tecido Muscularcarvalho30rebecaAinda não há avaliações
- Introdução à terapia pelo movimentoDocumento10 páginasIntrodução à terapia pelo movimentoKarina FerrazAinda não há avaliações
- Trabalho de Tecido Muscular.Documento2 páginasTrabalho de Tecido Muscular.elaideccamara100% (1)
- Sistema de SustentaçãoDocumento19 páginasSistema de SustentaçãoEder100% (1)
- Classificação Dos Tecidos, Relacionando As Suas Funções e Localização Dos Tecidos para A Formação Dos ÓrgãosDocumento26 páginasClassificação Dos Tecidos, Relacionando As Suas Funções e Localização Dos Tecidos para A Formação Dos ÓrgãosRenderson GonçalvesAinda não há avaliações
- Fundamentos da Ciência da CarneDocumento6 páginasFundamentos da Ciência da CarneDaniel BravoAinda não há avaliações
- O QUE REVISAR-Segunda Fase UVV-2022-1Documento2 páginasO QUE REVISAR-Segunda Fase UVV-2022-1Paulo BastosAinda não há avaliações
- Projeto Eletromicrografias para MOF142 2023 01Documento91 páginasProjeto Eletromicrografias para MOF142 2023 01Pablo MonticeliAinda não há avaliações
- Atividade Livro Todas As UnidDocumento19 páginasAtividade Livro Todas As UnidAnderson RibeiroAinda não há avaliações
- Aula 1 Vet - Introdução À Anatomia, Termos de PosiçãoDocumento98 páginasAula 1 Vet - Introdução À Anatomia, Termos de PosiçãoMARCELA PIMENTAAinda não há avaliações
- Resumo Da Prova de HistologiaDocumento5 páginasResumo Da Prova de Histologiaalle27Ainda não há avaliações
- Musculação Guto GalambaDocumento24 páginasMusculação Guto GalambaMiguel StrondaAinda não há avaliações
- Trabalho de recuperação ciências músculos e sentidosDocumento7 páginasTrabalho de recuperação ciências músculos e sentidosJéssica LayneAinda não há avaliações


























































![ESTRUTURA[1]..](https://imgv2-1-f.scribdassets.com/img/document/725364009/149x198/bec26b4e30/1713755014?v=1)