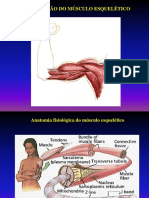
Você também pode gostar
- FisiologiaDocumento120 páginasFisiologiaMagno DuarteAinda não há avaliações
- HistologiaPL - Tecido MuscularDocumento35 páginasHistologiaPL - Tecido Muscularviolleta gomia100% (1)
- Tecido MuscularDocumento6 páginasTecido MuscularmayraAinda não há avaliações
- Problema 01 - Essa Fraqueza É GraveDocumento16 páginasProblema 01 - Essa Fraqueza É Gravematheus galvãoAinda não há avaliações
- Tecido MuscularDocumento9 páginasTecido MuscularIsadora RuthAinda não há avaliações
- Tecido Muscular - ResumoDocumento6 páginasTecido Muscular - ResumoHelena MarcondesAinda não há avaliações
- Apg s8p1Documento3 páginasApg s8p1gabrielAinda não há avaliações
- Sistema Muscular - FisiologiaDocumento5 páginasSistema Muscular - FisiologiaKauan GabrielAinda não há avaliações
- Resumo Tecido Muscular - JunqueiraDocumento4 páginasResumo Tecido Muscular - JunqueiraLucas Thiago BorgesAinda não há avaliações
- Contração Do Músculo EsqueléticoDocumento11 páginasContração Do Músculo EsqueléticoBeatriz Teixeira MonteiroAinda não há avaliações
- Contrao MuscularDocumento3 páginasContrao MuscularBreno Nunes da Silva OliveiraAinda não há avaliações
- FISIOLOGIA - RESUMO - Contração Do Musculo EsqueléticoDocumento2 páginasFISIOLOGIA - RESUMO - Contração Do Musculo EsqueléticoLara Geovana BezerraAinda não há avaliações
- OpenAccess Lorente 9788580393996 08Documento12 páginasOpenAccess Lorente 9788580393996 08Larissa Medicina veterináriaAinda não há avaliações
- Fisiologia MuscularDocumento17 páginasFisiologia MuscularMaria Baião BasílioAinda não há avaliações
- Apg 20 - Semestre 2Documento4 páginasApg 20 - Semestre 2Bruna BuenoAinda não há avaliações
- Junção NeuroDocumento33 páginasJunção NeuroFafnir Kesk100% (2)
- Fisiologia Da Contrac3a7c3a3o Muscular - RoDocumento36 páginasFisiologia Da Contrac3a7c3a3o Muscular - RoMarcelo CostaAinda não há avaliações
- Tecido MuscularDocumento6 páginasTecido MuscularLeo RamosAinda não há avaliações
- Estudo Dirigido 2Documento19 páginasEstudo Dirigido 2moematimoAinda não há avaliações
- Fechamento Tutoria 12Documento12 páginasFechamento Tutoria 12vanessa da silva caetanoAinda não há avaliações
- Aula 021659031827Documento12 páginasAula 021659031827Jurandi AraujoAinda não há avaliações
- Tecido MuscularDocumento4 páginasTecido MuscularDeadmanAinda não há avaliações
- Fisiologia Da Contração MuscularDocumento3 páginasFisiologia Da Contração MuscularVictor Carvalho NogueiraAinda não há avaliações
- FisiologiaDocumento38 páginasFisiologiaAleida SantosAinda não há avaliações
- Músculos e Receptores MuscularesDocumento6 páginasMúsculos e Receptores MuscularesLuciani MoreiraAinda não há avaliações
- 13 Tec MuscularDocumento47 páginas13 Tec MuscularmalubastosAinda não há avaliações
- Sistema MuscularDocumento18 páginasSistema MuscularGessica Fabiana Cardoso0% (1)
- Capitulo 6 Resumo Fisiologia GuytonDocumento5 páginasCapitulo 6 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- Contração Do Músculo EsqueléticoDocumento43 páginasContração Do Músculo Esqueléticorodrigo RodmorAinda não há avaliações
- Bioquímica Da Força e Contração MuscularDocumento10 páginasBioquímica Da Força e Contração MuscularhellensabinoAinda não há avaliações
- Medresumos 2016 - OmfDocumento16 páginasMedresumos 2016 - OmfMirianAinda não há avaliações
- Sistema Muscular EsqueléticoDocumento32 páginasSistema Muscular Esqueléticogoncalo camposAinda não há avaliações
- Biofísica Dos Sistemas Integradores: Prof. Romildo NogueiraDocumento19 páginasBiofísica Dos Sistemas Integradores: Prof. Romildo NogueiraThiago De Alencar LacerdaAinda não há avaliações
- ED. Fisio 16.09Documento6 páginasED. Fisio 16.09Nilo BarretoAinda não há avaliações
- Tecido MuscularDocumento3 páginasTecido MuscularCamilla Pereira100% (1)
- Tecido MuscularDocumento5 páginasTecido MusculargabrieltavaresAinda não há avaliações
- Contração Muscular - SarcômeroDocumento7 páginasContração Muscular - SarcômeroJoyceMedeirosAinda não há avaliações
- #Resumo#Documento10 páginas#Resumo#Monitoriadefisio100% (2)
- Fisiologia Da Contração Muscular EsqueléticaDocumento33 páginasFisiologia Da Contração Muscular EsqueléticaSara LimaAinda não há avaliações
- Tecido MuscularDocumento85 páginasTecido MuscularLarissa MendesAinda não há avaliações
- Tecido MuscularDocumento3 páginasTecido MuscularRaquel ChequeleiroAinda não há avaliações
- Fisiologia Da Contração Muscular - AulaDocumento6 páginasFisiologia Da Contração Muscular - AulaISADHORA MARIA MARAN DE SOUZAAinda não há avaliações
- Aula 4 - Estrutura e Função Dos Músculos - PPTX U Nirg LKDSJDocumento38 páginasAula 4 - Estrutura e Função Dos Músculos - PPTX U Nirg LKDSJBruna Roberta PimentaAinda não há avaliações
- Tecido MuscularDocumento7 páginasTecido MuscularRebeca SoaresAinda não há avaliações
- Fisiologia Músculo Estriado Esquelético 2023Documento36 páginasFisiologia Músculo Estriado Esquelético 2023Fabio Ferreira LorenaAinda não há avaliações
- Resumo Contração MuscularDocumento11 páginasResumo Contração MuscularThiago PontesAinda não há avaliações
- Tecido MuscularDocumento3 páginasTecido Muscularcedaspy cedaspyAinda não há avaliações
- Fisiologia Veterinaria IDocumento6 páginasFisiologia Veterinaria IMelyssa NacaratiAinda não há avaliações
- Trabalho Contração Muscular FinalDocumento37 páginasTrabalho Contração Muscular FinalNaillyne GalvãoAinda não há avaliações
- 1contração Do Músculo EsqueléticoDocumento63 páginas1contração Do Músculo EsqueléticoKelly Coelho RochaAinda não há avaliações
- Aula Contração MuscularDocumento21 páginasAula Contração MuscularLucas ReisAinda não há avaliações
- Tecido MuscularDocumento5 páginasTecido MuscularRodrigo SousaAinda não há avaliações
- Aula 2 - Contração Do Músculo Esquelético e LisoDocumento16 páginasAula 2 - Contração Do Músculo Esquelético e LisoMatheus Messina ScorsoliniAinda não há avaliações
- Resumo Tecido Muscular Histologia e Fisiologia Da ContraçãoDocumento10 páginasResumo Tecido Muscular Histologia e Fisiologia Da ContraçãoIgor BarbosaAinda não há avaliações
- Fala FisiologiaDocumento5 páginasFala FisiologiaRachel PassosAinda não há avaliações
- Resumo Tecido MuscularDocumento5 páginasResumo Tecido MuscularLaryssa AmorimAinda não há avaliações
- Tecido Muscular 2015Documento57 páginasTecido Muscular 2015Webert Martins MeloAinda não há avaliações
- Resenha Condromalacia PatelarDocumento2 páginasResenha Condromalacia PatelarThiago Oliveira de SousaAinda não há avaliações
- M10P2 - Sistema Motor SomáticoDocumento8 páginasM10P2 - Sistema Motor Somáticoana flavia furtado telesAinda não há avaliações
- Fisiologia Do Exercicio - Roger de Moraes - EstácioDocumento305 páginasFisiologia Do Exercicio - Roger de Moraes - EstácioCaio SilvaAinda não há avaliações
- Fisiologia e Neurofisiologia Da Micção ApostilaDocumento36 páginasFisiologia e Neurofisiologia Da Micção ApostilaMarques Davdson Santos100% (1)
- Mov Ameb Voo Ins BivDocumento27 páginasMov Ameb Voo Ins BivRoberto SantosAinda não há avaliações
- Lição Transformação Do Músculo em Carne, Influência Na Qualidade Da CarneDocumento84 páginasLição Transformação Do Músculo em Carne, Influência Na Qualidade Da CarneLuana CavalcanteAinda não há avaliações
- UrsulinoCarolinaMaccagnaniCampos TCCDocumento78 páginasUrsulinoCarolinaMaccagnaniCampos TCCAnnalethycia PereiraAinda não há avaliações
- Q05 - Questionário Semana 05 - 2021C - Bases Biológicas Aplicadas À Educação Física - Educação Física (53906)Documento6 páginasQ05 - Questionário Semana 05 - 2021C - Bases Biológicas Aplicadas À Educação Física - Educação Física (53906)Marcos Wiggy100% (1)
- Roteiro 1 - Tecido MuscularDocumento9 páginasRoteiro 1 - Tecido MusculardievellenAinda não há avaliações
- Alterações Pós MortaisDocumento12 páginasAlterações Pós MortaisMillena MartinsAinda não há avaliações
- Atividade Teleaula IiiDocumento3 páginasAtividade Teleaula IiiBinho MirandaAinda não há avaliações
- UFPR 2018 Anatovet 1 Aula 10 Miologia GERALDocumento53 páginasUFPR 2018 Anatovet 1 Aula 10 Miologia GERALMarcelo Hillk IPTV100% (1)
- Prática Viii - Soi IiDocumento12 páginasPrática Viii - Soi IiFabricio Rodrimaur Rodrigues GonçalvesAinda não há avaliações
- Biofísica Dos Sistemas Integradores: Prof. Romildo NogueiraDocumento19 páginasBiofísica Dos Sistemas Integradores: Prof. Romildo NogueiraThiago De Alencar LacerdaAinda não há avaliações
- Slide Musculo EsqueléticoDocumento54 páginasSlide Musculo EsqueléticoAdrya ThayanneAinda não há avaliações
- UP1Documento113 páginasUP1Ana Catarina MaiaAinda não há avaliações
- Unidade Motora e Suas SindromesDocumento39 páginasUnidade Motora e Suas SindromesCibele BragaAinda não há avaliações
- Suplementacao Na Atividade FisicaDocumento251 páginasSuplementacao Na Atividade FisicaGuilhermeAinda não há avaliações
- Sistema MuscularDocumento65 páginasSistema MuscularThelmaAinda não há avaliações
- Biomedicina EstéticaDocumento72 páginasBiomedicina Estéticajo.dorneles7Ainda não há avaliações
- Aula Musculo EsqueleticoDocumento37 páginasAula Musculo EsqueleticoFERNANDO VINÍCIUS ARAUJO DELMONDESAinda não há avaliações
- EMOV - M2 - Manual Do AlunoDocumento202 páginasEMOV - M2 - Manual Do AlunoFrancisco Martins100% (2)
- Querida MadrinhaDocumento44 páginasQuerida MadrinhaCefas AlmeidaAinda não há avaliações
- Fisiologia ResumoDocumento7 páginasFisiologia ResumoBianca SartiAinda não há avaliações
- Fechamento Tutoria 12Documento12 páginasFechamento Tutoria 12vanessa da silva caetanoAinda não há avaliações
- Exercícios de Revisão BiocelDocumento115 páginasExercícios de Revisão BiocelMaria Rosa DmengeonAinda não há avaliações
- 1bi - Modulo 2Documento8 páginas1bi - Modulo 2TaironeAinda não há avaliações
- Citoesqueleto PDFDocumento7 páginasCitoesqueleto PDFDiogo Mendonça100% (1)
- Módulo 3 - Caso 4 - ContraçõesDocumento16 páginasMódulo 3 - Caso 4 - ContraçõesEmilly GomesAinda não há avaliações
- Manual de Ergonomia - Adaptando o Trabalho Ao Homem - Etienne GrandjeanDocumento174 páginasManual de Ergonomia - Adaptando o Trabalho Ao Homem - Etienne GrandjeanRafAinda não há avaliações
- Ergonomia 5Documento16 páginasErgonomia 5Jane CastroAinda não há avaliações
- CMS Tegumentar, Reprodutor e Locomotor - Livro - U3Documento60 páginasCMS Tegumentar, Reprodutor e Locomotor - Livro - U3Eniberto AlissonAinda não há avaliações