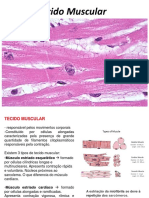
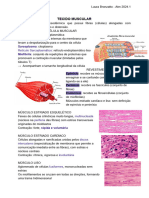
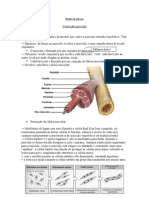
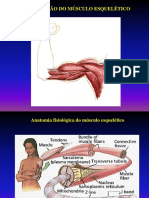
Você também pode gostar
- Contração muscular: mecanismo e fontes de energiaDocumento3 páginasContração muscular: mecanismo e fontes de energiaFelipe DiasAinda não há avaliações
- Contração muscularDocumento41 páginasContração muscularbut zinhoAinda não há avaliações
- Contração do Músculo EsqueléticoDocumento56 páginasContração do Músculo EsqueléticoMargareth CruvinelAinda não há avaliações
- Tecido Muscular - ResumoDocumento6 páginasTecido Muscular - ResumoHelena MarcondesAinda não há avaliações
- Capítulo 6 e 7Documento4 páginasCapítulo 6 e 7Lara Geovana BezerraAinda não há avaliações
- Capitulo 6 Resumo Fisiologia GuytonDocumento5 páginasCapitulo 6 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- Sistema Músculo-EsqueléticoDocumento21 páginasSistema Músculo-EsqueléticoiotticarolAinda não há avaliações
- Fisiologia Contraçao MuscularDocumento3 páginasFisiologia Contraçao MuscularEDUARDO NEVES CHIUCHIAinda não há avaliações
- Fisiologia: Músculo Estriado EsqueléticoDocumento5 páginasFisiologia: Músculo Estriado EsqueléticoCarolina DibAinda não há avaliações
- Arquitetura muscular e contraçãoDocumento120 páginasArquitetura muscular e contraçãoMagno DuarteAinda não há avaliações
- Potencial de Ação - Contração MuscularDocumento9 páginasPotencial de Ação - Contração MuscularRodrigo SousaAinda não há avaliações
- FisiologiaDocumento9 páginasFisiologiakarol oliosiAinda não há avaliações
- Gasto - Problema 4 - Tecido MuscularDocumento5 páginasGasto - Problema 4 - Tecido Musculargabriel pupoAinda não há avaliações
- Problema 01 - Essa Fraqueza É GraveDocumento16 páginasProblema 01 - Essa Fraqueza É Gravematheus galvãoAinda não há avaliações
- Contração MuscularDocumento7 páginasContração MuscularBeatriz Teixeira MonteiroAinda não há avaliações
- Sistema MuscularDocumento23 páginasSistema MuscularWarleyAinda não há avaliações
- Musculo EsqueléticoDocumento3 páginasMusculo Esqueléticosoneska wdymAinda não há avaliações
- Resenha Condromalacia PatelarDocumento2 páginasResenha Condromalacia PatelarThiago Oliveira de SousaAinda não há avaliações
- Movimento humano ATP músculos esqueléticos contraçãoDocumento7 páginasMovimento humano ATP músculos esqueléticos contraçãoJoyceMedeirosAinda não há avaliações
- Documentos JuniorDocumento9 páginasDocumentos Juniorresumos medAinda não há avaliações
- Resumo Tecido Muscular - JunqueiraDocumento4 páginasResumo Tecido Muscular - JunqueiraLucas Thiago BorgesAinda não há avaliações
- Bioquímica da contração muscular: filamentos deslizantesDocumento10 páginasBioquímica da contração muscular: filamentos deslizanteshellensabinoAinda não há avaliações
- Açao NeuromuscularDocumento53 páginasAçao Neuromuscularanon_736807922Ainda não há avaliações
- Fisiologia Veterinaria IDocumento6 páginasFisiologia Veterinaria IMelyssa NacaratiAinda não há avaliações
- APG 20 - Tecido MuscularDocumento13 páginasAPG 20 - Tecido Muscularlarac0706Ainda não há avaliações
- Fisiologia Av 2 k4zDocumento89 páginasFisiologia Av 2 k4zCaroline MoreiraAinda não há avaliações
- O Musculo CardiacoDocumento2 páginasO Musculo CardiacoCamilla JordãoAinda não há avaliações
- Contração muscular: estrutura, mecanismo e doençasDocumento63 páginasContração muscular: estrutura, mecanismo e doençasmark3116Ainda não há avaliações
- Músculo Esquelético e LisoDocumento9 páginasMúsculo Esquelético e LisoDudu VitaAinda não há avaliações
- Fisiologia Da Contrac3a7c3a3o Muscular - RoDocumento36 páginasFisiologia Da Contrac3a7c3a3o Muscular - RoMarcelo CostaAinda não há avaliações
- Contração muscular em 40Documento16 páginasContração muscular em 40Matheus Messina ScorsoliniAinda não há avaliações
- #Resumo#Documento10 páginas#Resumo#Monitoriadefisio100% (2)
- Apg s8p1Documento3 páginasApg s8p1gabrielAinda não há avaliações
- Fisiologia Da Contração Muscular EsqueléticaDocumento33 páginasFisiologia Da Contração Muscular EsqueléticaSara LimaAinda não há avaliações
- Tecido MuscularDocumento6 páginasTecido MuscularmayraAinda não há avaliações
- Contrao MuscularDocumento3 páginasContrao MuscularBreno Nunes da Silva OliveiraAinda não há avaliações
- 1bi - Modulo 2Documento8 páginas1bi - Modulo 2TaironeAinda não há avaliações
- Contração Do Músculo EsqueléticoDocumento6 páginasContração Do Músculo EsqueléticoJulia SallesAinda não há avaliações
- Contração do músculo esqueléticoDocumento37 páginasContração do músculo esqueléticoFERNANDO VINÍCIUS ARAUJO DELMONDESAinda não há avaliações
- Resumo Fisiologia MuscularDocumento6 páginasResumo Fisiologia MuscularDaniel SilvaAinda não há avaliações
- Medresumos 2016 - OmfDocumento16 páginasMedresumos 2016 - OmfMirianAinda não há avaliações
- Fisiologia Músculo EsqueléticoDocumento21 páginasFisiologia Músculo EsqueléticoLucas ReisAinda não há avaliações
- Contracao Do Musculo LisoDocumento5 páginasContracao Do Musculo LisoAna Clara GardioliAinda não há avaliações
- Tecido MuscularDocumento6 páginasTecido Muscularlau24bronzattoAinda não há avaliações
- Tecido MuscularDocumento4 páginasTecido MuscularDeadmanAinda não há avaliações
- Músculos e contração muscularDocumento36 páginasMúsculos e contração muscularmaysaAinda não há avaliações
- Introdução à fisiologia muscularDocumento25 páginasIntrodução à fisiologia muscularThatianny FernandesAinda não há avaliações
- Sistema Muscular - FisiologiaDocumento5 páginasSistema Muscular - FisiologiaKauan GabrielAinda não há avaliações
- ED. Fisio 16.09Documento6 páginasED. Fisio 16.09Nilo BarretoAinda não há avaliações
- Contração muscularDocumento6 páginasContração muscularMelissa100% (2)
- Fisiologia Da Contração Muscular - AulaDocumento6 páginasFisiologia Da Contração Muscular - AulaISADHORA MARIA MARAN DE SOUZAAinda não há avaliações
- Músculo esquelético: estrutura e contraçãoDocumento5 páginasMúsculo esquelético: estrutura e contraçãomariany100% (2)
- Contração Muscular - Biossistemas Do Corpo Humano (Leonardo) - 19 04 21Documento9 páginasContração Muscular - Biossistemas Do Corpo Humano (Leonardo) - 19 04 21Skcr DoorsKAinda não há avaliações
- Cabulas Dos Musculos 1Documento1 páginaCabulas Dos Musculos 1api-26429188Ainda não há avaliações
- Histologia - Junqueira & CarneiroDocumento22 páginasHistologia - Junqueira & CarneiroBeatriz GoulartAinda não há avaliações
- Aula 7 - Tecido MuscularDocumento32 páginasAula 7 - Tecido MuscularBrenda SoutoAinda não há avaliações
- Resumo Sobre As Proteínas MuscularesDocumento2 páginasResumo Sobre As Proteínas MuscularesGeovana BeatrizAinda não há avaliações
- 1contração Do Músculo EsqueléticoDocumento63 páginas1contração Do Músculo EsqueléticoKelly Coelho RochaAinda não há avaliações
- Fisiologia do músculo esqueléticoDocumento12 páginasFisiologia do músculo esqueléticoAna CostaAinda não há avaliações
- Apostila Ecg Versão 2018Documento12 páginasApostila Ecg Versão 2018Anonymous n4gtOaochAinda não há avaliações
- Drogas Vasoativas (NOVO)Documento4 páginasDrogas Vasoativas (NOVO)laris_rachelAinda não há avaliações
- Atividade 4 - Bio - Hematologia Clínica - 51-2023Documento4 páginasAtividade 4 - Bio - Hematologia Clínica - 51-2023Cavalini Assessoria AcadêmicaAinda não há avaliações
- Anatomia Coronária Com Angiografia Por TomografiaDocumento4 páginasAnatomia Coronária Com Angiografia Por TomografiaRComancheAinda não há avaliações
- Cirurgia Cardíaca: Tratamentos e ProcedimentosDocumento89 páginasCirurgia Cardíaca: Tratamentos e ProcedimentosElaine Santos100% (1)
- Apostila Do Curso Primeiro SocorroDocumento17 páginasApostila Do Curso Primeiro SocorroMag MagnocesarAinda não há avaliações
- Sistema cardiovascular: função, regulação e exercícioDocumento23 páginasSistema cardiovascular: função, regulação e exercícioNatália ValleAinda não há avaliações
- Os principais pontos na ventosaterapia: Ashi, gatilhos, tender e acupontosDocumento14 páginasOs principais pontos na ventosaterapia: Ashi, gatilhos, tender e acupontosgabrielAinda não há avaliações
- Aula Débito CardíacoDocumento35 páginasAula Débito CardíacoCarolina Yume ArazawaAinda não há avaliações
- Sistemas de transporte em animais sem circulaçãoDocumento5 páginasSistemas de transporte em animais sem circulaçãoBeatriz CostaAinda não há avaliações
- Hemostasia: os 4 eventos da coagulação sanguíneaDocumento12 páginasHemostasia: os 4 eventos da coagulação sanguíneaAthirson SousaAinda não há avaliações
- Farmacologia Do Sistema Nervoso AutônomoDocumento25 páginasFarmacologia Do Sistema Nervoso AutônomoMARCELO ANTONIO SILVA MENEZESAinda não há avaliações
- Funções e características dos principais tipos de leucócitosDocumento1 páginaFunções e características dos principais tipos de leucócitosCaroline Martins de AndradeAinda não há avaliações
- Infografico Sinais VitaisDocumento1 páginaInfografico Sinais VitaisInstituto Treni GestãoAinda não há avaliações
- Impressao Exames 16614685084036Documento2 páginasImpressao Exames 16614685084036André Luiz Reche de LimaAinda não há avaliações
- Tetralogia de FallotDocumento6 páginasTetralogia de FallotWalter SenaAinda não há avaliações
- ACLS Certification CourseDocumento36 páginasACLS Certification CourseROBSON PESSOAAinda não há avaliações
- Terminologia Da DorDocumento30 páginasTerminologia Da DorMarcelo Benedet Tournier100% (3)
- 5 Primeiros SocorrosDocumento75 páginas5 Primeiros SocorrosVALDECIR PEREIRA DE LIMAAinda não há avaliações
- Capítulo IV - Embolia, Infarto e ChoqueDocumento18 páginasCapítulo IV - Embolia, Infarto e ChoquePaula Canavezi de OliveiraAinda não há avaliações
- Apostila Cardiologia PDFDocumento92 páginasApostila Cardiologia PDFPedro HenriqueAinda não há avaliações
- Hemograma - Anemia e LeucemiasDocumento92 páginasHemograma - Anemia e LeucemiasJaqueline CarvalhoAinda não há avaliações
- Componentes do SangueDocumento24 páginasComponentes do SangueELIS CAMPOS WAKAMOTOAinda não há avaliações
- 514219ABORH01092023090325Documento13 páginas514219ABORH01092023090325kleberson arrudaAinda não há avaliações
- Controle da pressão arterial por receptoresDocumento2 páginasControle da pressão arterial por receptoresEdnardo RamosAinda não há avaliações
- Apostila Patologia GeralDocumento12 páginasApostila Patologia GeralGleici S2 Alves100% (1)
- Fisiologia cardiovascular resumoDocumento137 páginasFisiologia cardiovascular resumoVíctor MorelliAinda não há avaliações
- Circulacao Fetal e NeonatalDocumento15 páginasCirculacao Fetal e NeonatalKelson GomesAinda não há avaliações
- Você Sabe o Que É HomeostaseDocumento5 páginasVocê Sabe o Que É HomeostaseMilton AlencarAinda não há avaliações
- Metabolismo energético: respiração e fotossínteseDocumento6 páginasMetabolismo energético: respiração e fotossínteseEdy VianaAinda não há avaliações