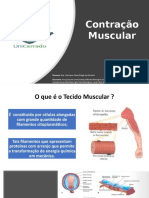
Você também pode gostar
- Mude A Sua Vida Com A Auto-Hipnose (PDFDrive)Documento160 páginasMude A Sua Vida Com A Auto-Hipnose (PDFDrive)semczukAinda não há avaliações
- FisiologiaDocumento120 páginasFisiologiaMagno DuarteAinda não há avaliações
- Neurociencias Parte 2Documento85 páginasNeurociencias Parte 2Alcione RodriguesAinda não há avaliações
- Guia Prático para Transtornos Emocionais - Ansiedade e Depressão Módulo IDocumento37 páginasGuia Prático para Transtornos Emocionais - Ansiedade e Depressão Módulo ICristina Fiorin CalegaroAinda não há avaliações
- Histologia Do Sistema NervosoDocumento60 páginasHistologia Do Sistema NervosoMax Nunes100% (1)
- ED. Fisio 16.09Documento6 páginasED. Fisio 16.09Nilo BarretoAinda não há avaliações
- Sistema MuscularDocumento43 páginasSistema MuscularIvanCostaPassosAinda não há avaliações
- Apostila 1 Sistema MuscularDocumento10 páginasApostila 1 Sistema MuscularSuellen OliveiraAinda não há avaliações
- Sistema NervosoDocumento81 páginasSistema NervosoGuilherme RebliAinda não há avaliações
- O Cérebro Nosso de Cada Dia - SinapsesDocumento3 páginasO Cérebro Nosso de Cada Dia - SinapsesJéssica Byron100% (1)
- Tecido MuscularDocumento3 páginasTecido Muscularcedaspy cedaspyAinda não há avaliações
- 20221110-17 - Módulo 4 - Sistema - Nervoso - e - Exercício - Fisiologia - Do - Exercício - I - PSS - 91 - SLIDESDocumento93 páginas20221110-17 - Módulo 4 - Sistema - Nervoso - e - Exercício - Fisiologia - Do - Exercício - I - PSS - 91 - SLIDESJoel MarquesAinda não há avaliações
- Sistema MuscularDocumento18 páginasSistema MuscularGessica Fabiana Cardoso0% (1)
- Histologia Tecido MuscularDocumento5 páginasHistologia Tecido Muscularmariany100% (2)
- Anatomia e Fisiologia Humana PDFDocumento46 páginasAnatomia e Fisiologia Humana PDFEdi PaivaAinda não há avaliações
- ARTIGO - Fisiologia Do Músculo EsqueléticoDocumento32 páginasARTIGO - Fisiologia Do Músculo EsqueléticoKeyla SilvaAinda não há avaliações
- Junção NeuroDocumento33 páginasJunção NeuroFafnir Kesk100% (2)
- Contração Muscular - SarcômeroDocumento7 páginasContração Muscular - SarcômeroJoyceMedeirosAinda não há avaliações
- Bioquímica Da Força e Contração MuscularDocumento10 páginasBioquímica Da Força e Contração MuscularhellensabinoAinda não há avaliações
- Medresumos 2016 - OmfDocumento16 páginasMedresumos 2016 - OmfMirianAinda não há avaliações
- Fisiologia Do Sistema MuscularDocumento17 páginasFisiologia Do Sistema MuscularKaillany Andrade NaivaAinda não há avaliações
- Resumo Contração MuscularDocumento11 páginasResumo Contração MuscularThiago PontesAinda não há avaliações
- Contração MuscularDocumento7 páginasContração MuscularBeatriz Teixeira MonteiroAinda não há avaliações
- Tecidos Muscular e NervosoDocumento17 páginasTecidos Muscular e NervosoBreno RiveroAinda não há avaliações
- Fisiologia Da Contração MuscularDocumento3 páginasFisiologia Da Contração MuscularVictor Carvalho NogueiraAinda não há avaliações
- Aula 2 - Contração Do Músculo Esquelético e LisoDocumento16 páginasAula 2 - Contração Do Músculo Esquelético e LisoMatheus Messina ScorsoliniAinda não há avaliações
- #Resumo#Documento10 páginas#Resumo#Monitoriadefisio100% (2)
- Fisiologia Do Musc EsqueléticoDocumento32 páginasFisiologia Do Musc EsqueléticosalvadoorjrAinda não há avaliações
- Sistema Músculo-EsqueléticoDocumento21 páginasSistema Músculo-EsqueléticoiotticarolAinda não há avaliações
- Contração Muscular Fisiologia Da Contração Muscular É o Espasmo MMMMMMMMMMMMMMMMDocumento12 páginasContração Muscular Fisiologia Da Contração Muscular É o Espasmo MMMMMMMMMMMMMMMMdelmindaAinda não há avaliações
- 02 - Estrutura Do MúsculoDocumento38 páginas02 - Estrutura Do MúsculorosicleriAinda não há avaliações
- Resumo Tecido Muscular - JunqueiraDocumento4 páginasResumo Tecido Muscular - JunqueiraLucas Thiago BorgesAinda não há avaliações
- Fechamento Tutoria 12Documento12 páginasFechamento Tutoria 12vanessa da silva caetanoAinda não há avaliações
- Tecido Muscular 2015Documento57 páginasTecido Muscular 2015Webert Martins MeloAinda não há avaliações
- Sistema MuscularDocumento7 páginasSistema MuscularCaroline TavaresAinda não há avaliações
- Tecido MuscularDocumento3 páginasTecido MuscularRaquel ChequeleiroAinda não há avaliações
- Fisiologia Av 2 k4zDocumento89 páginasFisiologia Av 2 k4zCaroline MoreiraAinda não há avaliações
- Ergonomia 5Documento16 páginasErgonomia 5Jane CastroAinda não há avaliações
- Anatomia General Dos Musculos-1Documento13 páginasAnatomia General Dos Musculos-1martagumbauAinda não há avaliações
- Músculos e Receptores MuscularesDocumento6 páginasMúsculos e Receptores MuscularesLuciani MoreiraAinda não há avaliações
- Ergonomia 5Documento16 páginasErgonomia 5Nicolle HenriquesAinda não há avaliações
- Tecido MuscularDocumento4 páginasTecido MuscularDeadmanAinda não há avaliações
- (9,0) Prova 1 - Fisiologia de SextaDocumento7 páginas(9,0) Prova 1 - Fisiologia de SextaLuis Felipe TeixeiraAinda não há avaliações
- FisiologiaDocumento9 páginasFisiologiakarol oliosiAinda não há avaliações
- Trabalho Contração Muscular FinalDocumento37 páginasTrabalho Contração Muscular FinalNaillyne GalvãoAinda não há avaliações
- Capitulo 8 Reumo Fisiologia GuytonDocumento4 páginasCapitulo 8 Reumo Fisiologia Guytonmuller maia100% (1)
- Fala FisiologiaDocumento5 páginasFala FisiologiaRachel PassosAinda não há avaliações
- Tecido MuscularDocumento7 páginasTecido MuscularRebeca SoaresAinda não há avaliações
- Anatomofisiologia I 2 FreqDocumento103 páginasAnatomofisiologia I 2 FreqAna MoreiraAinda não há avaliações
- Sistema Muscular - FisiologiaDocumento5 páginasSistema Muscular - FisiologiaKauan GabrielAinda não há avaliações
- Fisiologia Basica Aula 5Documento14 páginasFisiologia Basica Aula 5Karla MoniqueAinda não há avaliações
- Resumo Fisiologia MuscularDocumento6 páginasResumo Fisiologia MuscularDaniel SilvaAinda não há avaliações
- Fisio Juncal NeuroDocumento14 páginasFisio Juncal NeuroMaria Júlia CruzAinda não há avaliações
- Anatomia e Fisiologia Do Sistema MuscularDocumento27 páginasAnatomia e Fisiologia Do Sistema MuscularAdão 21Ainda não há avaliações
- Sistema Muscular - Material de ApoioDocumento5 páginasSistema Muscular - Material de ApoioOzimar FroesAinda não há avaliações
- Resumo - Módulo 1 - Fisiologia Do ExercícioDocumento23 páginasResumo - Módulo 1 - Fisiologia Do ExercícioLarissa PammelaAinda não há avaliações
- Sistema MuscularDocumento24 páginasSistema MuscularGabrielly SouzaAinda não há avaliações
- Fisiologia Da Contrac3a7c3a3o Muscular - RoDocumento36 páginasFisiologia Da Contrac3a7c3a3o Muscular - RoMarcelo CostaAinda não há avaliações
- Aula 4 - Estrutura e Função Dos Músculos - PPTX U Nirg LKDSJDocumento38 páginasAula 4 - Estrutura e Função Dos Músculos - PPTX U Nirg LKDSJBruna Roberta PimentaAinda não há avaliações
- RESUMO - Contração MuscularDocumento3 páginasRESUMO - Contração MuscularIngrid DomingosAinda não há avaliações
- Fisiologia - Musculo EsqueléticoDocumento12 páginasFisiologia - Musculo EsqueléticoHumberto KabukiAinda não há avaliações
- OpenAccess Lorente 9788580393996 08Documento12 páginasOpenAccess Lorente 9788580393996 08Larissa Medicina veterináriaAinda não há avaliações
- Tecido MuscularDocumento9 páginasTecido MuscularIsadora RuthAinda não há avaliações
- FISIOLOGIA MUSCULAR - CompressedDocumento73 páginasFISIOLOGIA MUSCULAR - CompressedcodigoderesgatedochaptersAinda não há avaliações
- Contra CPDFDocumento49 páginasContra CPDFGabriel VianaAinda não há avaliações
- Temática - Ciências da Vida - Relação e CoordenaçãoNo EverandTemática - Ciências da Vida - Relação e CoordenaçãoAinda não há avaliações
- FTG 5 Guião - 1.1. O CérebroDocumento9 páginasFTG 5 Guião - 1.1. O CérebrocarmoAinda não há avaliações
- Arquivos Simbidor 2007Documento404 páginasArquivos Simbidor 2007Rachel Marini ChioteAinda não há avaliações
- Disfuncao e Teste de MitchellDocumento31 páginasDisfuncao e Teste de MitchellfabioAinda não há avaliações
- Transporte Transmembranar e Propagação Do Impulso NervosoDocumento13 páginasTransporte Transmembranar e Propagação Do Impulso NervosoFilipa Vieira100% (2)
- Avaliação Primeiro Ano Faustino 4 BimestreDocumento2 páginasAvaliação Primeiro Ano Faustino 4 BimestreGracy PachecoAinda não há avaliações
- Memoria de Trabalho e Função ExecutivaDocumento79 páginasMemoria de Trabalho e Função ExecutivaAbel Albuquerque100% (1)
- Sistema EndocanabinoidesDocumento8 páginasSistema EndocanabinoidesAnuska ErikaAinda não há avaliações
- Potencial de Ação e SinapsesDocumento9 páginasPotencial de Ação e SinapsesAmanda Francisquini100% (1)
- P MunapsysDocumento28 páginasP MunapsysInstituto Ricco PortoAinda não há avaliações
- Papel Da Noradrenalina Na CogniçãoDocumento20 páginasPapel Da Noradrenalina Na Cogniçãolesca_S2Ainda não há avaliações
- Seminário Toxoplasmose PDFDocumento20 páginasSeminário Toxoplasmose PDFThaís DurãesAinda não há avaliações
- Junção NeuromuscularDocumento5 páginasJunção NeuromuscularRenildo da CruzAinda não há avaliações
- FISIOLOGIADocumento114 páginasFISIOLOGIAGiulia MannesAinda não há avaliações
- Prova Biologia 2020 MIMVDocumento15 páginasProva Biologia 2020 MIMVCatarinaFerreiraAinda não há avaliações
- Rafaela Monografia Versao FinalDocumento49 páginasRafaela Monografia Versao FinalMellina LisboaAinda não há avaliações
- Ação Dos Inseticidas Nos InsetosDocumento36 páginasAção Dos Inseticidas Nos InsetosEvandro KellerAinda não há avaliações
- Explorando A Mente Humana - e Book - 20240311 - 204732 - 0000Documento65 páginasExplorando A Mente Humana - e Book - 20240311 - 204732 - 0000mari.gimenes06Ainda não há avaliações
- 1 - Bases Anatômicas e Fisiológicas Do MovimentoDocumento7 páginas1 - Bases Anatômicas e Fisiológicas Do MovimentomktAinda não há avaliações
- Sinalização NeuronalDocumento10 páginasSinalização NeuronalSara MarquesAinda não há avaliações
- Neurofisiologia - ResumoDocumento78 páginasNeurofisiologia - Resumofejare2171Ainda não há avaliações
- Apontamentos PsicofisiologiaDocumento69 páginasApontamentos PsicofisiologiaiolandadiasmAinda não há avaliações
- Introdução Às Redes Neurais Artificiais - Prof Jorge Barreto UFSC PDFDocumento57 páginasIntrodução Às Redes Neurais Artificiais - Prof Jorge Barreto UFSC PDFvitor_pedroAinda não há avaliações