Você também pode gostar
- Aula08BioqI Funà à oProteà Nas PDFDocumento35 páginasAula08BioqI Funà à oProteà Nas PDFAbdul Laura CondulaAinda não há avaliações
- Hemoglobina e MioglobinaDocumento45 páginasHemoglobina e MioglobinaMiguel TuriniAinda não há avaliações
- Função ProteicaDocumento56 páginasFunção ProteicaGisele MeganAinda não há avaliações
- Resumo Funções de ProteínasDocumento6 páginasResumo Funções de ProteínasHeloisa Moreira Dias PereiraAinda não há avaliações
- Aminoácidos, Pepitideos e ProteinasDocumento36 páginasAminoácidos, Pepitideos e ProteinasRafael CamposAinda não há avaliações
- Bioinorgnica 1 - Fontes de Energia e FunesDocumento49 páginasBioinorgnica 1 - Fontes de Energia e FunesNathália CarvalhoAinda não há avaliações
- Aula 4 - Ligações QuímicasDocumento18 páginasAula 4 - Ligações QuímicasBaptistaAinda não há avaliações
- 3 Função Proteica Não Catalítica e EnzimasDocumento107 páginas3 Função Proteica Não Catalítica e EnzimasLuiza TavaresAinda não há avaliações
- 5 Funcao - De.proteinasDocumento25 páginas5 Funcao - De.proteinasJoão Vitor Souza de OliveiraAinda não há avaliações
- 2 Série - QuímicaDocumento6 páginas2 Série - QuímicaANDRE LUIZ RODRIGUES DA SILVAAinda não há avaliações
- Reações de Substituição Nucleofílica Ao Carbono SaturadoDocumento36 páginasReações de Substituição Nucleofílica Ao Carbono Saturadoprof.amanluisaAinda não há avaliações
- Epage - Pub - 5 Acidez e Basicidade de Compostos OrganicosDocumento55 páginasEpage - Pub - 5 Acidez e Basicidade de Compostos OrganicosAssis GomesAinda não há avaliações
- Reações de Substituição Nucleofílica Alifática Ao Carbono SaturadoDocumento30 páginasReações de Substituição Nucleofílica Alifática Ao Carbono SaturadoDaniel SilveiraAinda não há avaliações
- QO-Cap.10-SISTEMAS INSATURADOS CONJUGADOS - Resumo-2012Documento23 páginasQO-Cap.10-SISTEMAS INSATURADOS CONJUGADOS - Resumo-2012Margarida MirandaAinda não há avaliações
- AULA 4 - DienoDocumento49 páginasAULA 4 - DienoTaciana GuarnieriAinda não há avaliações
- Introdução As Reações Organicas Aula 01Documento32 páginasIntrodução As Reações Organicas Aula 01Natalia LopesAinda não há avaliações
- Os 5 Assuntos de Química Mais Cobrados No ENEMDocumento15 páginasOs 5 Assuntos de Química Mais Cobrados No ENEMPriscilla BritoAinda não há avaliações
- Biofisica Atomos Ions e MoleculasDocumento7 páginasBiofisica Atomos Ions e MoleculasManuel Baptista ManuelAinda não há avaliações
- Tom 1Documento134 páginasTom 1Josiel HenryAinda não há avaliações
- AT18 FosforilacaoOxidativaDocumento33 páginasAT18 FosforilacaoOxidativaMateus GarciaAinda não há avaliações
- Aula - 4 - Geometria Molecular e Forças IntermolecularesDocumento29 páginasAula - 4 - Geometria Molecular e Forças Intermolecularesra280421Ainda não há avaliações
- Bioenergética e MetabolismoDocumento5 páginasBioenergética e MetabolismoSamuel LemosAinda não há avaliações
- Aula - 4 - Geometria Molecular e Forças IntermolecularesDocumento29 páginasAula - 4 - Geometria Molecular e Forças IntermolecularesJuliano Augustus100% (1)
- Aula 03 Geom, Polar e ForçDocumento29 páginasAula 03 Geom, Polar e ForçLussandra Marquez MeneghelAinda não há avaliações
- Aula 3 - Teoria de Huckel - 20182Documento22 páginasAula 3 - Teoria de Huckel - 20182Jennifer Kupas RamosAinda não há avaliações
- Aula 5 - Ligações Químicas Parte IIDocumento60 páginasAula 5 - Ligações Químicas Parte IIGustavo Carvalho SilvaAinda não há avaliações
- Arilson 07Documento5 páginasArilson 07Juliane GrasielaAinda não há avaliações
- Aula 4 - Interações IntermolecularesDocumento64 páginasAula 4 - Interações IntermolecularesJulia HondaAinda não há avaliações
- Cadeia RespiratóriaDocumento60 páginasCadeia RespiratóriaAlline RodriguesAinda não há avaliações
- Ligações Químicas e ÁguaDocumento32 páginasLigações Químicas e ÁguaEmerson FranciscoAinda não há avaliações
- Chemistry BondDocumento62 páginasChemistry BondpriscielensilvestreAinda não há avaliações
- Artigo - SÍNTESE E CARACTERIZAÇÃO DO BIS (DIFENILFOSFINO) METANODocumento9 páginasArtigo - SÍNTESE E CARACTERIZAÇÃO DO BIS (DIFENILFOSFINO) METANOdpaulo79100% (2)
- Fluxo Transmembrana 2017 1Documento20 páginasFluxo Transmembrana 2017 1Carlos FelipeAinda não há avaliações
- Lig Quim Lig Ionica e TRPECV Elenir 2022 Engenha - 220924 - 104105Documento52 páginasLig Quim Lig Ionica e TRPECV Elenir 2022 Engenha - 220924 - 104105Jeisy SilveiraAinda não há avaliações
- Tom 1 PDFDocumento42 páginasTom 1 PDFmanuellaamattos19Ainda não há avaliações
- Relatório Referente Ao Dia 02 - 09Documento18 páginasRelatório Referente Ao Dia 02 - 09Carolina Beatriz MarinhoAinda não há avaliações
- 02 - Proteinas GlobularesDocumento42 páginas02 - Proteinas GlobularesIgor FortunatiAinda não há avaliações
- CadeiaRespiratoria FosforilacaoOxidativa (2022 I)Documento36 páginasCadeiaRespiratoria FosforilacaoOxidativa (2022 I)Gustavo ErbertAinda não há avaliações
- Fosforilação OxidativaDocumento36 páginasFosforilação OxidativaCatharina GuimarãesAinda não há avaliações
- Aula HemoglobinasDocumento32 páginasAula HemoglobinasSergio S SouzaAinda não há avaliações
- Haletos de AlquiloDocumento26 páginasHaletos de AlquiloIsraele CostaAinda não há avaliações
- Ligacoes SecundariasDocumento22 páginasLigacoes Secundariasbrunofc86Ainda não há avaliações
- Propriedades e Estruturas Dos Compostos QuímicosDocumento11 páginasPropriedades e Estruturas Dos Compostos QuímicosJonesM CraftAinda não há avaliações
- Apontamentos - Bioinorgânica PDFDocumento14 páginasApontamentos - Bioinorgânica PDFFCiênciasAinda não há avaliações
- Trabalho de BioquímicaDocumento5 páginasTrabalho de BioquímicaJéssica AlmeidaAinda não há avaliações
- Lista QuimicaDocumento13 páginasLista QuimicaLohana Rodrigues PereiraAinda não há avaliações
- 5 - Ligações QuímicasDocumento28 páginas5 - Ligações QuímicasAchilles BermudesAinda não há avaliações
- Aula 14Documento38 páginasAula 14le.lima742Ainda não há avaliações
- 06 Proteinas - Funcoes (2 Por Pagina)Documento19 páginas06 Proteinas - Funcoes (2 Por Pagina)Stella Indiukov dos SantosAinda não há avaliações
- Obtenção de Matéria Pelos Seres AutotróficosDocumento8 páginasObtenção de Matéria Pelos Seres AutotróficosAndreia GonçalvesAinda não há avaliações
- Turma Enem-IFPI-Geometria-Polaridade-Ligações Intermoleculares.Documento18 páginasTurma Enem-IFPI-Geometria-Polaridade-Ligações Intermoleculares.Janielson VianaAinda não há avaliações
- Resumo - Cadeia de Transporte de Elétrons e Fosforilação OxidativaDocumento5 páginasResumo - Cadeia de Transporte de Elétrons e Fosforilação OxidativaLAIS VERONEZ MARQUES BARROSAinda não há avaliações
- Capítulo 13 - Princípios de BioenergéticaDocumento3 páginasCapítulo 13 - Princípios de BioenergéticaJean Carlos Araujo BrilhanteAinda não há avaliações
- Eletroqu MicaDocumento7 páginasEletroqu MicaMário FilhoAinda não há avaliações
- Aula - 3 - Ligações QuímicasDocumento45 páginasAula - 3 - Ligações QuímicasJuliano AugustusAinda não há avaliações
- Ácidos e Bases de Brönsted e Lowry: Uma visão aplicada ao meio ambienteNo EverandÁcidos e Bases de Brönsted e Lowry: Uma visão aplicada ao meio ambienteAinda não há avaliações
- Apostila Química Orgânica: Carbono, Dienos E AromáticosNo EverandApostila Química Orgânica: Carbono, Dienos E AromáticosAinda não há avaliações
- Biofísica para ciências biomédicas – 4ª ediçãoNo EverandBiofísica para ciências biomédicas – 4ª ediçãoAinda não há avaliações
- Aula 1 - Logística e Cadeia de SuprimentosDocumento37 páginasAula 1 - Logística e Cadeia de SuprimentosEMANUEL_CHAVESAinda não há avaliações
- Modelo de Relatorio Anual de AtividadesDocumento6 páginasModelo de Relatorio Anual de AtividadesNascimento Passos de OliveiraAinda não há avaliações
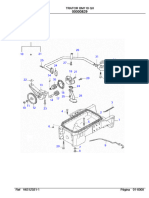
- Trator Trator Bm110 Bm110 Gii GIIDocumento2 páginasTrator Trator Bm110 Bm110 Gii GIIWashington SAinda não há avaliações
- Apostila Trato Gastrointestinal IIIDocumento29 páginasApostila Trato Gastrointestinal IIIDenise VerganiAinda não há avaliações
- Como Produzir Conservas Que Não Estragam - Antonino PaixãoDocumento64 páginasComo Produzir Conservas Que Não Estragam - Antonino PaixãoJosemar SilvaAinda não há avaliações
- PH 03Documento19 páginasPH 03MafefefesinhaAinda não há avaliações
- E.r.9 Ano v3 - Cad 02.super - EnsinoDocumento22 páginasE.r.9 Ano v3 - Cad 02.super - EnsinoROSENILSON RODRIGUESAinda não há avaliações
- Comunicação Nas EmpresasDocumento31 páginasComunicação Nas EmpresasAline Landim25% (4)
- Gabarito - Volume 09Documento10 páginasGabarito - Volume 09gabytottidulliusAinda não há avaliações
- Habilidades para Uma Carreira de Sucesso Na EngenhariaDocumento5 páginasHabilidades para Uma Carreira de Sucesso Na EngenhariaAna Julia BicalhoAinda não há avaliações
- Limitações Dos Aparelhos Nucleares No Controlo de Qualidade de AterrosDocumento11 páginasLimitações Dos Aparelhos Nucleares No Controlo de Qualidade de AterrosZemar Jose ManuelAinda não há avaliações
- 03 As Origens Da Psicoterapia de Crianças e AdolescentesDocumento43 páginas03 As Origens Da Psicoterapia de Crianças e AdolescentesEAD100% (1)
- Relatório de Estágio Obrigatório - Marcus v. de B. SilvaDocumento59 páginasRelatório de Estágio Obrigatório - Marcus v. de B. SilvaVinícius de BritoAinda não há avaliações
- Ficha Informativa de Entidade Não ContábilDocumento1 páginaFicha Informativa de Entidade Não ContábilThariny22Ainda não há avaliações
- Jacques MaritainDocumento3 páginasJacques MaritainGladius100% (1)
- CP 14 Metodos de EnergiaDocumento77 páginasCP 14 Metodos de Energiaatila diasAinda não há avaliações
- Avaliação Autismo em AdultoDocumento4 páginasAvaliação Autismo em AdultoAdriana Rubio WodewotzkiAinda não há avaliações
- Fagifor. FortalezaDocumento61 páginasFagifor. FortalezaMécia costaAinda não há avaliações
- Folder MedilabDocumento12 páginasFolder MedilabHerick SavioneAinda não há avaliações
- Catalogo CorfioDocumento28 páginasCatalogo CorfioJoão MenegazAinda não há avaliações
- Avaliacao Neuropsicologica Da LinguagemDocumento11 páginasAvaliacao Neuropsicologica Da LinguagemJAILENE PIRESAinda não há avaliações
- Artigo - Produção de Conteúdo para WebDocumento3 páginasArtigo - Produção de Conteúdo para WebMatheus HipólitoAinda não há avaliações
- Apostila Saúde OcupacionalDocumento200 páginasApostila Saúde OcupacionalBruno MelloAinda não há avaliações
- E-Book GRADIENT BOOSTINGDocumento36 páginasE-Book GRADIENT BOOSTINGDiogo Lima BarretoAinda não há avaliações
- Certikin CTX2011Documento302 páginasCertikin CTX2011ruipatrick1Ainda não há avaliações
- Combinação Fluticasona e Azelastina Intranasal No Tratamento de Adolescentes Com Rinite Alérgica de Difícil ControleDocumento8 páginasCombinação Fluticasona e Azelastina Intranasal No Tratamento de Adolescentes Com Rinite Alérgica de Difícil ControleDaniel TolentinoAinda não há avaliações
- Primeira Lista 2021-01Documento2 páginasPrimeira Lista 2021-01Rodrigo MartinsAinda não há avaliações
- Modelo de Relatório Do Estágio CurricularDocumento11 páginasModelo de Relatório Do Estágio CurricularVictória SousaAinda não há avaliações
- Biometria Aplicada Ao Estudo Da Diversidade GenéticaDocumento626 páginasBiometria Aplicada Ao Estudo Da Diversidade Genética28rhq6mbfkAinda não há avaliações
- TCC - Linux - Rodrigo Dall AlbaDocumento61 páginasTCC - Linux - Rodrigo Dall Albardallalba100% (1)