Você também pode gostar
- Relacionamento Hospedeiro MicrorganismoDocumento7 páginasRelacionamento Hospedeiro MicrorganismoAndreia LuísAinda não há avaliações
- Microbiologia aplicada à enfermagemDocumento17 páginasMicrobiologia aplicada à enfermagemsandramadeira-1Ainda não há avaliações
- Aspectos Ecológicos Da Relação Bactéria Hospedeiro e Biofilmes CBDocumento51 páginasAspectos Ecológicos Da Relação Bactéria Hospedeiro e Biofilmes CBMarcelo Partes de OliveiraAinda não há avaliações
- Mecanismos Patogenese BacterianaDocumento22 páginasMecanismos Patogenese BacterianaAndreia OliveiraAinda não há avaliações
- Ecossistemas microbiológicosDocumento12 páginasEcossistemas microbiológicosEmeEme ImportaçõesAinda não há avaliações
- INTERAÇÃO MICRORGANISMO X HOSPEDEIRODocumento36 páginasINTERAÇÃO MICRORGANISMO X HOSPEDEIROadestradora.erikaAinda não há avaliações
- Interações parasita-hospedeiroDocumento5 páginasInterações parasita-hospedeiromaria eduardaAinda não há avaliações
- Apostila de Microbiologia MédicaDocumento64 páginasApostila de Microbiologia MédicaElizabeth Honorina LouroAinda não há avaliações
- Microbiota anfibiôntica e relações hospedeiro-bactériaDocumento64 páginasMicrobiota anfibiôntica e relações hospedeiro-bactériaRosanne VenturaAinda não há avaliações
- Microbiota anfibiôntica e relações hospedeiro-bactériaDocumento64 páginasMicrobiota anfibiôntica e relações hospedeiro-bactériaTony Jefferson100% (2)
- Microbiologia Médica I: Patógenos e Microbioma HumanoNo EverandMicrobiologia Médica I: Patógenos e Microbioma HumanoAinda não há avaliações
- PEPARAR AULA!!! Patogênese-Das-Doenças-Bacterianas-20181Documento49 páginasPEPARAR AULA!!! Patogênese-Das-Doenças-Bacterianas-20181Marcella Manfredi100% (2)
- Patogênese Das Doenças Bacterianas 0000Documento49 páginasPatogênese Das Doenças Bacterianas 0000Rogerio CeniAinda não há avaliações
- Riscos Biológicos PDFDocumento28 páginasRiscos Biológicos PDFWaldean SantanaAinda não há avaliações
- Infetocontagiosas - 2 Aula - Catarina Pinto - ESAI 2023Documento62 páginasInfetocontagiosas - 2 Aula - Catarina Pinto - ESAI 2023RitaAinda não há avaliações
- M. Madalena AlmeidaDocumento8 páginasM. Madalena AlmeidaMadalena FaléAinda não há avaliações
- Doenças Infecciosas GuiaDocumento24 páginasDoenças Infecciosas GuiaVictor HugoAinda não há avaliações
- microbiotanormaleIRAS 20220407173039Documento16 páginasmicrobiotanormaleIRAS 20220407173039Luana NevesAinda não há avaliações
- Sistema imune e microrganismosDocumento30 páginasSistema imune e microrganismosCASA DA VÓAinda não há avaliações
- Mecanismo de Resistencia Do HospedeiroDocumento7 páginasMecanismo de Resistencia Do HospedeiroEdirlenesaraAinda não há avaliações
- Aula 01 - MicrobiologiaDocumento54 páginasAula 01 - Microbiologianeto100% (1)
- Introdução à ParasitologiaDocumento7 páginasIntrodução à ParasitologiaMárcio MendesAinda não há avaliações
- 3 Aula Microbiologia - Oral - Resistência - Do - OrganismoDocumento35 páginas3 Aula Microbiologia - Oral - Resistência - Do - OrganismoJefferson PradoAinda não há avaliações
- 05 Imunidade, Vacinas e Soros (Aula 01)Documento68 páginas05 Imunidade, Vacinas e Soros (Aula 01)Nathália do Vale100% (1)
- Microbiota Normal e Patógenos BacterianosDocumento13 páginasMicrobiota Normal e Patógenos BacterianospedroAinda não há avaliações
- Resumo Fatores de Virulência Nos FungosDocumento8 páginasResumo Fatores de Virulência Nos FungosRafaele Rodrigues100% (7)
- Microrganismos e doenças infecciosasDocumento37 páginasMicrorganismos e doenças infecciosasVictor MafraAinda não há avaliações
- Patogenia Da Infecção Bacteriana e Resistê Ncia Do Hospedeiro À InfecçãoDocumento27 páginasPatogenia Da Infecção Bacteriana e Resistê Ncia Do Hospedeiro À Infecçãomandy_diiAinda não há avaliações
- Fatores Ambientais Como Determinantes de DoençaDocumento6 páginasFatores Ambientais Como Determinantes de DoençaAmanda BeatrizAinda não há avaliações
- Introdução às Doenças Infecciosas: Agentes, Imunidade e FatoresDocumento12 páginasIntrodução às Doenças Infecciosas: Agentes, Imunidade e FatoresManjuma Afonso CarreiroAinda não há avaliações
- Imuno 1Documento28 páginasImuno 1Jonatan Dos Santos De OliveiraAinda não há avaliações
- Resumo LPF - Módulo CDocumento14 páginasResumo LPF - Módulo Cgabriella.ferreira.inacioAinda não há avaliações
- Microbiologia: Introdução aos microrganismosDocumento53 páginasMicrobiologia: Introdução aos microrganismosPamella de sousaAinda não há avaliações
- Patogenia Das Bactã©rias 22Documento7 páginasPatogenia Das Bactã©rias 22Angelique ViannyAinda não há avaliações
- Aula Microbiologia 1 ENFERMAGEM - IntroduçãoDocumento37 páginasAula Microbiologia 1 ENFERMAGEM - IntroduçãoIara RemediosAinda não há avaliações
- Introdução A Imunologia BucalDocumento3 páginasIntrodução A Imunologia BucalAirexTVAinda não há avaliações
- Doenças Infecciosas - Prova 1Documento19 páginasDoenças Infecciosas - Prova 1Rebeca Moreira Batista rmb2Ainda não há avaliações
- Microbiota humana e interações com microrganismosDocumento36 páginasMicrobiota humana e interações com microrganismosNatália LopesAinda não há avaliações
- Patologia pratica parasitasDocumento7 páginasPatologia pratica parasitasAna Luiza MenezesAinda não há avaliações
- Introdução à microbiologia e parasitologia: microrganismos, transmissão e prevençãoDocumento44 páginasIntrodução à microbiologia e parasitologia: microrganismos, transmissão e prevençãoDébora SantosAinda não há avaliações
- Resumo de Microbiologia - Taxonomia Bacteriana e Cadeia de InfecçãoDocumento12 páginasResumo de Microbiologia - Taxonomia Bacteriana e Cadeia de InfecçãoNataliaAinda não há avaliações
- Agentes biológicos: riscos e medidas de controleDocumento166 páginasAgentes biológicos: riscos e medidas de controlekarla moniqueAinda não há avaliações
- Patogenicidade e VirulênciaDocumento10 páginasPatogenicidade e VirulênciaAiuba ConstantinoAinda não há avaliações
- MicróbiosDocumento16 páginasMicróbiosCecília GuiseAinda não há avaliações
- Perguntas para EstudarDocumento12 páginasPerguntas para Estudarthais camposAinda não há avaliações
- Microbiologia e Parasitologia AULA 2 (1)Documento12 páginasMicrobiologia e Parasitologia AULA 2 (1)rayssaoliveiira05Ainda não há avaliações
- Sistema Imunológico - Vacinas - Ciências - Prof. JoãoDocumento27 páginasSistema Imunológico - Vacinas - Ciências - Prof. JoãoEdivon OliveiraAinda não há avaliações
- Agentes BiológicosDocumento43 páginasAgentes Biológicosluis_carlos_467009100% (2)
- 2.defesa Do OrganismoDocumento45 páginas2.defesa Do OrganismoPCris AlvesAinda não há avaliações
- Imunidade Inata - Cap 2Documento13 páginasImunidade Inata - Cap 2Natália ChiesaAinda não há avaliações
- 7 - MicrobiologiaDocumento22 páginas7 - MicrobiologiaImerson Alberto AlfaneAinda não há avaliações
- TAP5-cadeia EpidemiológicaDocumento6 páginasTAP5-cadeia EpidemiológicaAna Rita RainhoAinda não há avaliações
- CONCEITOS GERAIS SOBRE DOENÇAS INFECCIOSASDocumento6 páginasCONCEITOS GERAIS SOBRE DOENÇAS INFECCIOSASBernardo AugustoAinda não há avaliações
- Micro Biolog I ADocumento7 páginasMicro Biolog I Alarissagabriela.limaAinda não há avaliações
- CAV-T-EAD V - Aula 1.1 A 1.2 - Fundamentos de BiossegurançaDocumento6 páginasCAV-T-EAD V - Aula 1.1 A 1.2 - Fundamentos de BiossegurançaÁlex PaixãoAinda não há avaliações
- MicologiaDocumento11 páginasMicologiaMarcos FelipeAinda não há avaliações
- Isso Explicaria Seus Sintomas?: Entendendo os Riscos de Uma Epidemia Oculta da Doença de Lyme no BrasilNo EverandIsso Explicaria Seus Sintomas?: Entendendo os Riscos de Uma Epidemia Oculta da Doença de Lyme no BrasilAinda não há avaliações
- Tipos Sanguíneos, Transfusão e ReaçõesDocumento3 páginasTipos Sanguíneos, Transfusão e ReaçõesLuíza Ferreira De FigueiredoAinda não há avaliações
- Continuação Da Lista de Exercícios 1Documento3 páginasContinuação Da Lista de Exercícios 1adrianegs50% (2)
- Bacterias Parte IDocumento27 páginasBacterias Parte ILuan HenriqueAinda não há avaliações
- Revisão de Imunologia BásicaDocumento13 páginasRevisão de Imunologia Básicam.lauraborba04Ainda não há avaliações
- DSTDocumento2 páginasDSTRuth MorenoAinda não há avaliações
- Biologia CelulaDocumento7 páginasBiologia CelulaRafael Abreu CostaAinda não há avaliações
- Imunologia – ciência experimentalDocumento8 páginasImunologia – ciência experimentalnayaraAinda não há avaliações
- Caso Clinico 03 - MHC IDocumento3 páginasCaso Clinico 03 - MHC IJaqueline Ferraz RegoAinda não há avaliações
- Reino Monera e classificação de bactériasDocumento14 páginasReino Monera e classificação de bactériasGreicy MartinsAinda não há avaliações
- Vacinas e InjetaveisDocumento260 páginasVacinas e InjetaveisJaneFinn100% (2)
- Aula 6. Classificacao EucariotosDocumento28 páginasAula 6. Classificacao EucariotosAlexandre NazarethAinda não há avaliações
- 3 - Taxonomia e Gêneros de BactériasDocumento64 páginas3 - Taxonomia e Gêneros de BactériasSaulo Saturnino SouzaAinda não há avaliações
- Disbiose intestinal idosos probióticos prebióticosDocumento8 páginasDisbiose intestinal idosos probióticos prebióticosLucélia Alves Paixão100% (1)
- Morfologia e estrutura procarionteDocumento59 páginasMorfologia e estrutura procarionteSuilan Moreira FerreiraAinda não há avaliações
- Microbiologia AplicadaDocumento6 páginasMicrobiologia AplicadavcuryAinda não há avaliações
- Estudo Da Prova Base ComportamentoDocumento69 páginasEstudo Da Prova Base ComportamentomarefranteixeiraAinda não há avaliações
- Resistência bacteriana e principais espéciesDocumento5 páginasResistência bacteriana e principais espéciesAracy Helena MarquesAinda não há avaliações
- Biologia Celular - Aula 02Documento49 páginasBiologia Celular - Aula 02Hppfbbbw HppfbbbwAinda não há avaliações
- Mononucleose Infecciosa SlidesDocumento35 páginasMononucleose Infecciosa Slideskadydel cordovaAinda não há avaliações
- PneumoniaDocumento10 páginasPneumoniaMariaAinda não há avaliações
- Desidroquinato Desidratase (EC 4.2.1.10) e Chiquimato Desidrogenase (EC 1.1.1.25)Documento85 páginasDesidroquinato Desidratase (EC 4.2.1.10) e Chiquimato Desidrogenase (EC 1.1.1.25)Phablo vinicius dos santos carneiroAinda não há avaliações
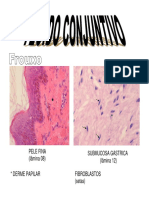
- Cap.2 - Tecido ConjuntivoDocumento13 páginasCap.2 - Tecido ConjuntivoLuana AguiarAinda não há avaliações
- Questões de Biologia - Genética MolecularDocumento15 páginasQuestões de Biologia - Genética Molecularnetoalvirubro100% (2)
- Atividade Genetica - 9 AnoDocumento3 páginasAtividade Genetica - 9 Anosabrina martinsAinda não há avaliações
- Genética - Resumo e Conceitos Básicos - Toda MatériaDocumento7 páginasGenética - Resumo e Conceitos Básicos - Toda MatériaCésar NardelloAinda não há avaliações
- Introdução Imunologia Aula 01Documento33 páginasIntrodução Imunologia Aula 01Marianne SilvaAinda não há avaliações
- 18 19 4 O Sangue PDFDocumento27 páginas18 19 4 O Sangue PDFAfonso CunhaAinda não há avaliações
- Funções das células T CD4+ efetoras na eliminação de microrganismosDocumento9 páginasFunções das células T CD4+ efetoras na eliminação de microrganismosFelipe GonzalezAinda não há avaliações
- Cromossomo SexualDocumento3 páginasCromossomo SexualraahrafsAinda não há avaliações
- Microbiologia - ProtozoáriosDocumento5 páginasMicrobiologia - ProtozoáriosJezielAinda não há avaliações