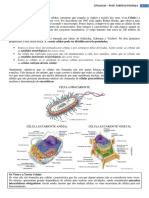
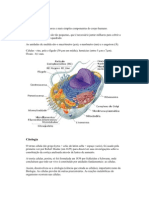
Você também pode gostar
- Questionário I - Biologia CelularDocumento6 páginasQuestionário I - Biologia CelularBianca Sa100% (6)
- Organelas e estruturas celularesDocumento2 páginasOrganelas e estruturas celularesjasmimangaAinda não há avaliações
- Introdução à Citologia - Organelas e Transporte CelularDocumento3 páginasIntrodução à Citologia - Organelas e Transporte Celularfátima castroAinda não há avaliações
- Anatomia e fisiologia do corpo humanoDocumento19 páginasAnatomia e fisiologia do corpo humanoFrancilene ModestoAinda não há avaliações
- Bases Biológicas Celulares e TecidaisDocumento14 páginasBases Biológicas Celulares e TecidaisMaria MariaAinda não há avaliações
- Biologia CelularDocumento7 páginasBiologia Celularwilkley.silvaAinda não há avaliações
- Respostas aos exercícios de biologia EJADocumento6 páginasRespostas aos exercícios de biologia EJASantannaServilAinda não há avaliações
- Histologia AplicadaDocumento27 páginasHistologia AplicadaKemuel AsafeAinda não há avaliações
- CitologiaDocumento11 páginasCitologiaSlúvio RapuluaAinda não há avaliações
- Primeira Atividade - Portfolio 1Documento7 páginasPrimeira Atividade - Portfolio 1Naiara SouzaAinda não há avaliações
- 12 Classe BiologiaDocumento55 páginas12 Classe BiologiaNelsonAngeloZameia92% (24)
- Embriologia, Citologia e HistologiaDocumento7 páginasEmbriologia, Citologia e HistologiaIvan MoreiraAinda não há avaliações
- Apostila de Fisiologia MédicaDocumento76 páginasApostila de Fisiologia MédicaMarco Antonio VianaAinda não há avaliações
- J6 UfbddpDocumento4 páginasJ6 UfbddpNanikaGNAinda não há avaliações
- O ESTUDO DAS CÉLULASDocumento5 páginasO ESTUDO DAS CÉLULASMT CRAFTAinda não há avaliações
- Biologia - Citologia - Profa KaireDocumento4 páginasBiologia - Citologia - Profa KaireKaire NardiAinda não há avaliações
- HISTOLOGIADocumento41 páginasHISTOLOGIAcarlosnalandaAinda não há avaliações
- Resumo Histologia JunqueiraDocumento8 páginasResumo Histologia JunqueiraEmilly EduardaAinda não há avaliações
- Sau001 LP Biologia Basica Aula2Documento12 páginasSau001 LP Biologia Basica Aula2Rafaela Sayuri Andrade GarciaAinda não há avaliações
- Diferenciação Parte I - HistologiaDocumento39 páginasDiferenciação Parte I - Histologiaivan_lalandaAinda não há avaliações
- CURSO DE EDUCAÇÃO FÍSICADocumento6 páginasCURSO DE EDUCAÇÃO FÍSICAGrazi MoreiraAinda não há avaliações
- Aula 1 Visão Geral Da CélulaDocumento33 páginasAula 1 Visão Geral Da Célulaapi-19738142Ainda não há avaliações
- Capítulo 7 - CelulasDocumento50 páginasCapítulo 7 - CelulasferreiraericaluanaAinda não há avaliações
- Introdução à Histologia VeterináriaDocumento81 páginasIntrodução à Histologia VeterináriaCarla Nogueira0% (1)
- Citologia - Estudo das célulasDocumento11 páginasCitologia - Estudo das célulasBeto AgostinhoAinda não há avaliações
- Aula Bocel 1Documento115 páginasAula Bocel 1loja xeroxAinda não há avaliações
- Características e estruturas dos seres vivos à escala celularDocumento28 páginasCaracterísticas e estruturas dos seres vivos à escala celularPaula FernandesAinda não há avaliações
- Revisão Bibliográfica UESPIDocumento9 páginasRevisão Bibliográfica UESPICarla Sousa SilvaAinda não há avaliações
- Modelo de Relatório - NUT Professora Vanessa MoreiraDocumento6 páginasModelo de Relatório - NUT Professora Vanessa Moreirakananda.s.subrinhoAinda não há avaliações
- Modelo de Relatório - NUT Professora Vanessa Moreira - Docx Celula AnimalDocumento6 páginasModelo de Relatório - NUT Professora Vanessa Moreira - Docx Celula Animalkananda.s.subrinhoAinda não há avaliações
- Biologia 12a A2Documento12 páginasBiologia 12a A2Moises FerroAinda não há avaliações
- A Base Celular Da VidaDocumento14 páginasA Base Celular Da VidaCarolina RodriguesAinda não há avaliações
- Citologia: Introdução aos principais componentes e teorias celularesDocumento11 páginasCitologia: Introdução aos principais componentes e teorias celularesIsaac VictorAinda não há avaliações
- 5 BiologiaDocumento114 páginas5 BiologiaLeo FAinda não há avaliações
- zinho 3 biologia celularDocumento14 páginaszinho 3 biologia celulararsenioAinda não há avaliações
- Conhecendo as células: estrutura e funçõesDocumento27 páginasConhecendo as células: estrutura e funçõesIzamarAinda não há avaliações
- Semana 3 - LabMorfo e MedLabDocumento8 páginasSemana 3 - LabMorfo e MedLabluisadurelloAinda não há avaliações
- Aula 3 - Biologia CelularDocumento18 páginasAula 3 - Biologia CelularcleidefersilvaAinda não há avaliações
- Biologia Celular IntroduçãoDocumento38 páginasBiologia Celular IntroduçãoMaxwell M. RibeiroAinda não há avaliações
- O conceito e componentes da célulaDocumento8 páginasO conceito e componentes da célulaRoberto Eduardo VieiraAinda não há avaliações
- Introdução Ao Estudo Da CélulaDocumento25 páginasIntrodução Ao Estudo Da Céluladrifrancas88100% (10)
- Relatorio AnimalDocumento5 páginasRelatorio Animalsaleskauanny535Ainda não há avaliações
- Respostas Estudo Dirigido 1 (Com Figurinhas)Documento17 páginasRespostas Estudo Dirigido 1 (Com Figurinhas)RogerMeloAinda não há avaliações
- Diversidadenabiosfera RFDocumento5 páginasDiversidadenabiosfera RFMarta FerrinhoAinda não há avaliações
- 3-CitologiaDocumento9 páginas3-CitologiaFelix Fernando LitoAinda não há avaliações
- Resumo HistologiaDocumento6 páginasResumo HistologiaAndressa Araújo100% (1)
- Célula AnimalDocumento6 páginasCélula AnimalFelipe MeloAinda não há avaliações
- Citologia - o Que É, Teoria Celular, Organelas e MaisDocumento19 páginasCitologia - o Que É, Teoria Celular, Organelas e Maisgm51topzeraAinda não há avaliações
- Sebenta Da 10 ClasseDocumento52 páginasSebenta Da 10 ClassedayanesouseAinda não há avaliações
- Aula 1Documento16 páginasAula 1Larissa Nacif CanalAinda não há avaliações
- Fisiopatologia Da Pele: Prof Beatriz Essenfelder BorgesDocumento97 páginasFisiopatologia Da Pele: Prof Beatriz Essenfelder Borgeslii pereiraAinda não há avaliações
- Organelas celularesDocumento10 páginasOrganelas celularesJoão LucasAinda não há avaliações
- Análise Da Citologia Histologia e FisiologiaDocumento25 páginasAnálise Da Citologia Histologia e FisiologiaStefânia MoraisAinda não há avaliações
- Ciências Da Natureza e Suas Tecnologias Ens. MédioDocumento72 páginasCiências Da Natureza e Suas Tecnologias Ens. MédioÉder MalletAinda não há avaliações
- Resumo de Histologia - Gênese e Desenvolvimento - Tecido Epitelial-1Documento31 páginasResumo de Histologia - Gênese e Desenvolvimento - Tecido Epitelial-1Renan SoaresAinda não há avaliações
- Sistema muscularDocumento38 páginasSistema muscularLeandro MartinsAinda não há avaliações
- Ross Tecido EpitelialDocumento71 páginasRoss Tecido EpitelialMaria Eduarda SantosAinda não há avaliações
- BIOMEMBRANASDocumento4 páginasBIOMEMBRANASaninhayomeAinda não há avaliações
- Anatomia e Histologia Dentária I FinalDocumento83 páginasAnatomia e Histologia Dentária I FinalMariana Gilvaz100% (1)
- Características Gerais Da Célula e Suas EstruturasDocumento29 páginasCaracterísticas Gerais Da Célula e Suas Estruturascerqueira gonzagaAinda não há avaliações
- Estrutura e funções do núcleo celularDocumento2 páginasEstrutura e funções do núcleo celularArthur CândidoAinda não há avaliações
- Plano de Aula - Histologia e Embriologia para OdontologiaDocumento6 páginasPlano de Aula - Histologia e Embriologia para OdontologiaimmesodontoAinda não há avaliações
- Estudo Dirigido Sistema CirculatórioDocumento10 páginasEstudo Dirigido Sistema CirculatóriokievnnyAinda não há avaliações
- Atividade de Aprendizagem. MuscularDocumento17 páginasAtividade de Aprendizagem. Muscularmarcosbigandrade89% (9)
- Técnicas de visualização de microrganismosDocumento12 páginasTécnicas de visualização de microrganismosjoao silva pintoAinda não há avaliações
- Análise Hematológica CompletaDocumento7 páginasAnálise Hematológica CompletaFabio PogianAinda não há avaliações
- Exercícios de Fixação - 3 - Organelas Celulares - 2023Documento5 páginasExercícios de Fixação - 3 - Organelas Celulares - 2023Esmeralda MuchisseAinda não há avaliações
- Pele e Anexos: Estrutura e FunçõesDocumento40 páginasPele e Anexos: Estrutura e FunçõesMarco Thúlio50% (2)
- APOSTILA de Célula VegetalDocumento12 páginasAPOSTILA de Célula VegetalKaliene Da Silva Carvalho Martins100% (1)
- Atlas - Biologia Dos TecidosDocumento14 páginasAtlas - Biologia Dos TecidosEvellyn SouzaAinda não há avaliações
- Técnicas HistológicasDocumento7 páginasTécnicas HistológicasLAYLA CLARELLIS SANTOS MOTA PINTOAinda não há avaliações
- Tecidos conjuntivo e adiposoDocumento25 páginasTecidos conjuntivo e adiposoMatheus de FreitasAinda não há avaliações
- Atividade 3 de Projeto IntegradorDocumento9 páginasAtividade 3 de Projeto Integradorvitor.vmAinda não há avaliações
- Divisão Celular: Mitose e MeioseDocumento4 páginasDivisão Celular: Mitose e MeioseELAINE CONCEIÇÃO ROCHAAinda não há avaliações
- AV3 - 9º Ano - 2º BimDocumento4 páginasAV3 - 9º Ano - 2º BimLeticia CardosoAinda não há avaliações
- Epitélio: estrutura, classificação e funções emDocumento15 páginasEpitélio: estrutura, classificação e funções emAstria DiasAinda não há avaliações
- AULA 6 Mitose - MeioseDocumento54 páginasAULA 6 Mitose - MeioseGabriel Fonseca SantosAinda não há avaliações
- Atividade2 - Organelas - Citoplasmáticas - e - Suas - Funções - Bio - 2EMDocumento3 páginasAtividade2 - Organelas - Citoplasmáticas - e - Suas - Funções - Bio - 2EMFlavia GouveiaAinda não há avaliações
- Ficha de Exercicios Anatomia PDFDocumento4 páginasFicha de Exercicios Anatomia PDFjuga diqueAinda não há avaliações
- Controle Neuromuscular Do MovimentoDocumento45 páginasControle Neuromuscular Do MovimentoNavegantes Silva de OliveiraAinda não há avaliações
- Portfólio II Biologia HumanaDocumento5 páginasPortfólio II Biologia HumanaJuliano Gonçalves de OliveiraAinda não há avaliações
- Trabalho ABNT PolitécnicoDocumento21 páginasTrabalho ABNT PolitécnicoMarcia PiresAinda não há avaliações
- Resumo de CitoplasmaDocumento2 páginasResumo de CitoplasmaAna Laura Campos de PieriAinda não há avaliações
- Transporte de proteínas para organelasDocumento8 páginasTransporte de proteínas para organelasJéssica CarvalhoAinda não há avaliações
- HistologiaDocumento19 páginasHistologiaCarol MoraesAinda não há avaliações